2016-01-06 17:05:22
УЗИ – добровольная платная мутация – расплата через 15-20 лет В нашей стране УЗИ появилось с ...
+ развернуть текстсохранённая копия
УЗИ – добровольная платная мутация – расплата через 15-20 лет В нашей стране УЗИ появилось с разрушением контроля системы защиты… далее →
2015-03-11 05:15:35
... информации. Пётр Гаряев переворачивает наши представления ... компьютер. Пётр Гаряев+ развернуть текстсохранённая копия
Наш организм использует технологии телепортации информации. Пётр Гаряев переворачивает наши представления о строении человеческого организма. Генетический код человека оперирует понятием нуля. Текст ДНК и РНК является произведением разумного существа. Как выглядит это разумное существо? ДНК излучает торсионные поля и звуки. Генетический аппарат человека – квантовый биокомпьютер. Клетки нашего организма знакомы друг с другом и общаются … Читать далее Достояние Планеты: Человек – Это квантовый компьютер. Пётр Гаряев→
Прионы – класс видоспецифичных паразитических белков нескольких линий (штаммов), вызывающих нейродистрофии у животных и людей [18]. Cтэнли Прузинер, получивший за исследования в этой области Нобелевскую премию 1997г., назвал эти белки в начале 80-х прионами (protein infectious virion). Прионы вызывают заболевания типа “скрэйпи” (у овец) и так называемого “коровьего бешенства” или губчатого энцефалита. У людей – это болезни “куру”, “синдром Крейцфельда-Якоба”, “синдром Гертсмана-Штраусслера-Шейнкера”, “синдром Альперса”, “фатальная семейная инсомния”. В этой области достигнуты большие успехи. И все-таки, ключевые тонкие механизмы развития патологических состояний такого рода остаются неизвестными [23]. Мыши с нокаутом гена prn-p устойчивы к инфекции PrPsc, что демонстрирует необходимость наличия PrPc для развития губчатой энцефалопатии [8]. Были изолировали РНК-аптамеры (РА), которые могли точно распознавать рекомбинантный прионовый белок хомяка, присоединенный к глутатион S-трансферазе. При этом РА были чувствительны к определенным аминокислотным последовательностям. Характерным для РА является наличие гуанин-обогащенного изгиба РНК с образованием локальных 4-х спиральных участков РНК с повторяющимися гуаниновыми квартетами, которые названы Г-квадриплексами или Г-тетрадами. Существенно, что они характерны и для хромосомных теломер. Такая 4-х спиральность проявляется как неканоническое спаривание оснований, отличное от Уотсон-Криковского и существующее благодаря стабилизации такого комплекса аминокислотами белков [23]. В цитируемой работе Вейса и др. были поставлены вопросы, имеющие некоторое отношение к предлагаемой идее работы генома на иных принципах (см. ниже). Эти вопросы таковы: (i) как РА распознают прионовые белки?, (ii) как РА связываются с прионовыми белками? (iii) достаточна ли точность распознавания РА-ми прионовых белков в гомогенатах мозга инфицированных и неифицированных животных?
Прионовые белки (PrPsc) имеют штаммовую специфичность, и в этом они подобны бактериям и вирусам. Фенотип и функции последних определяются их геномами. Но прионы, если их выделить из пораженных тканей в чистом виде, не имеют в своем составе нуклеиновых кислот. Попадая в желудок (или другие ткани), прионы мигрируют каким-то необъяснимым образом в головной мозг, размножаются там, вызывая его морфо-функциональную деградацию. Непонятно как преодолевается дистанция Желудок Мозг, перекрытая гематоэнцефалическим барьером. Это остается загадкой, хотя есть предположения, что лимфа служит промежуточным этапом транспорта PrPsc в головной мозг. Предполагается, что этот белок может через нервные окончания ретроградно проникать по аксонам в спинной и головной мозг. Последний механизм, хотя не объяснен и не доказан, имеет определенное обоснование в новой теории нервного импульса [25] (см. ниже). В организме людей, животных, и даже дрожжей синтезируются нормальные не инфекционные прионоподобные белки (PrPc), аналогичные по своей аминокислотной последовательности белкам прионов. Имеются и соответствующие гены, ответственные за синтез PrPc. Кроме отсутствия функции патогенности, PrPc отличаются от PrPsc по вторичной структуре. При их взаимодействии в мозгу или in vitro PrPc переходит в PrPsc с уменьшением доли -спиралей и увеличением области пептида, занятой -складчатой структурой. Все последующие порции синтезируемых в мозгу PrPc также приобретают - структуру и, соответственно, функцию PrPsc. Роль "нормального" PrPc в биосистемах остается пока неизвестной, хотя есть предположения, что она заключается в обеспечении нормального функционирования клеток Пуркинье [19].
Что касается дрожжевых прионов (Psi+ и Sup35) у Saccharamyces cerevisiae, то они поставили в тупик генетиков, поскольку оказалось, что они передают генетические наследуемые признаки без участия ДНК или РНК [University of Chicago Medical Center press release, 1997].
Самое непонятное, и с нашей точки зрения ключевое, в понимании сущности прионов - факт вирусоподобной штаммовой специфичности в патогенезе, вызываемом разными типами PrPsc (их известно более 20), при видимом отсутствии у них ДНК или РНК, то есть генетического аппарата. Гены различных PrPc незначительно отличаются по нуклеотидным последовательностям. Мутации этих генов могут вызывать конверсию PrPcPrPsc с последующим накоплением PrPsc и заболеванием. Известны случаи спонтанного образования прионовых штаммов у пожилых людей и старых животных. И это также непонятно. Размножение прионов длительно. Для мышей, в зависимости от штамма, продолжительность инкубационного периода от 50 до 500 дней. У людей это годы. Развитие прионов сопровождается макроскопическим, несовместимым с жизнью, накоплением в мозгу полимерных нитей PrPsc, которые способны к окрашиванию Конго красным и двойному лучепреломлению. Последнее означает, что прионовые бляшки вызывают расхождение лучей право- и лево поляризованного света. Этот, казалось бы незначительный факт, и вроде бы не имеющий отношения к прионовым синдромам, представляется не случайным по отношению к патогенным свойствам прионов (см. ниже).
ТЕЛОМЕРЫ И ТЕЛОМЕРАЗА
В последнее время резко усилился интерес к теломерам и теломеразе в аспекте проблемы старения (см., например, Биохимия, т.62, выпуск 11, ноябрь 1997г.; том целиком посвящен проблеме теломер и теломеразе). Последующее цитирование по теломерам взято из этого источника.
В 1961г. Л.Хейфлик и П.Мурхед показали ограниченность репликативной способности нормальных фибробластов человека. Когда нормальные эмбриональные клетки человека растут в наиболее благоприятных условиях, старение и смерть их неизбежно наступает после ~50 удвоений популяции. Это проявление внутренних свойств самих клеток. Наблюдение было воспроизведено другими многочисленными исследованиями. В то же время раковые клетки, пассируемые в аналогичных "идеальных" условиях бессмертны. В чем причина смертности одних клеток и бессмертия других? В 1971г. А.М.Оловников предположил, что причина старения и смерти на клеточном уровне заключается в недорепликации ДНК-полимеразой (теломеразой) концов хромосом (теломер) при клеточных делениях [31]. Это связано с использованием затравочных РНК-праймеров при синтезе ДНК от 5'-конца к 3'-концу и их последующим удалением. При этом 5'-конец реплики остается недореплицированным. С каждым актом репликации хромосом их концы укорачиваются на размер, занимаемый теломеразой. Этот размер иногда - своеобразная "мертвая зона", в которой не происходит удвоение однотяжных ДНК при делениях клеток. И это происходит до тех пор, пока не начинаются утраты жизненно важных кодирующих последовательностей ДНК, граничащих с теломерами. Такие утраты некоторые исследователи считают синонимом старения. Число укорочений теломер служит репликометром, определяющим количество делений, которые должна совершить нормальная клетка. После достижения минимального критического числа повторяющихся теломерных последовательностей TTAGGG, клетки теряют способность к делению. Так считалось до недавнего времени.
Оказалось, однако, что ситуация значительно сложнее. Существуют механизмы противостояния эффекту "мертвых зон". Один из них обнаружили Грейдер и Блэкберн на Tetrahymena. Именно эти исследователи открыли терминальную трансферазу - рибонуклеопротеиновый фермент, который получил название "теломераза". Оказалось, что теломеры после каждого деления клеток синтезируются теломеразой заново. Фермент достраивает 3' - конец теломер и, таким способом, удлиняет теломеры, компенсируя эффект "мертвой зоны", иногда с лихвой. Теломераза оказалась необычной обратной транскриптазой, то есть РНК зависимой ДНК полимеразой со своей собственной матрицей РНК для синтеза коротких повторяющихся последовательностей концевых ДНК хромосом. Наиболее хорошо изучена матричная область РНК Tetrahymena thermophila. Эта область содержит 9 нуклеотидных остатков в позиции от 43 до 51 (5'-CAACCCCAA-3'). Из них только 7 нуклеотидных остатков (43-49) являются собственно матричными, они составляют активную часть теломеразы и определяют каталитические функции фермента. Теломераза была затем обнаружена в экстрактах иммортализованных клеток человека. В отличие от нормальных смертных клеточных штаммов линии аномальных бессмертных клеток не стареют и продуцируют теломеразу. Поэтому теломеры иммортализованных клеток не укорачиваются при последовательных пассажах in vitro. Особенно эффективно такая защита от укорочений ДНК представлена у раковых клеток. В норме также обнаруживаются аналогичные процессы, например, в тканях плода и семенниках.
Есть еще одна особенность в механизмах сохранения теломер, которая, как и в случае синтеза прионов в головном мозгу в варианте болезни куру, непонятна и является предметом анализа настоящей статьи. Как уже ясно, иммортализация клеток человека в культуре обычно связана с экспрессией теломеразной активности. Однако, в некоторых случаях теломеразная активность не выявляется, хотя сравнение паттернов терминального рестрикционного фрагмента (TRF) до и после иммортализации показывает, что удлинение теломер действительно произошло. Крайняя гетерогенность длин теломер и различия динамики поддержания теломер в теломеразонегативных линиях в сравнении с теломеразопозитивными линиями показывают, что эти клетки использовали один или более альтернативных (ALT) механизмов удлинения теломер (ALT - Alternative Mechanism for Lengthening of Telomeres). Существенно, что все исследованные к настоящему времени линии клеток с ALT имеют сходный паттерн TRF. Это говорит, возможно, за общий механизм ALT. Все теломеразонегативные иммортализованные линии клеток, исследованные до настоящего времени, имели признаки ALT-активности, что согласуется с гипотезой, что поддержание теломер при помощи теломеразы или ALT необходимо для иммортализации. Природа механизма (или механизмов) ALT в настоящее время неизвестна, хотя есть предположение, не основанное на эксперименте, что здесь может работать механизм рекомбинационного удлинения теломер.
Таким образом, приходится констатировать, что в рамках ALT-пути имеет место необычный феномен синтеза ДНК "в отсутствие" кодирующей вещественной комплементарной матрицы ДНК или РНК. Это дополняет список сходных "аномалий", начатых с непонятных способа проникновения прионов в мозг и отчетливого вирусоподобного поведения прионовых белков при видимом отсутствии у них ДНК или РНК, то есть, в последнем случае, без геноструктур сохраняется информация о генетических штаммовых признаках прионов. Суда же попадает "аномалия" с дрожжевыми прионами, некоторые генетические признаки которых передаются опять-таки без матриц ДНК или РНК.
QB-РЕПЛИКАЗА
И, наконец, третий феномен, входящий в рассматриваемое семейство "безматричных аномалий" при синтезе полинуклеотидов. Это функционирование Qb-репликазы, РНК зависимой РНК-полимеразы колифага Qb. В системе in vitro Qb-репликаза может работать как машина саморепликации молекул РНК. Это показано достаточно давно [16, 20]. Замечательным оказалось свойство этого фермента синтезировать определенные последовательности коротких РНК без матрицы РНК [21]. Недавно аналогичный "безматричный" синтез РНК показан и для РНК полимеразы бактериофага T7 [5]. Такой же результат получен для синтеза РНК de novo посредством ДНК-зависимой РНК-полимеразы фагов Т7, Т3 и SP6 [Интернет]. В этих экспериментах снова нарушается центральная догма молекулярной биологии и генетики, постулирующая казалось бы незыблемое: ДНКРНКБЕЛОК. Это каноническое утверждение, что матрицами для синтеза ДНК или РНК могут служить только вещественные молекулы РНК или ДНК. Но в одном пункте эта догма уже модифицирована. Поток стратегической информации, как уже хорошо известно после открытия обратной транскриптазы, выглядит по другому: ДНКРНКБЕЛОК. Если учитывать "аномалии" при размножении прионов, то вполне возможна и еще одна поправка: ДНКРНКБЕЛОК, что будет обсуждаться ниже.
Поскольку "безматричный" синтез РНК приводит к необходимости дальнейших изменений в понимании работы генетического аппарата (по крайней мере, для низших биосистем) в научной литературе началась и продолжается обширная дискуссия о корректности "безматричных" экспериментов. Предельно высокая чистота опытов с Qb репликазой в отношении артефактов, связанных с наличием в реактивах и лабораторной посуде посторонних следовых примесей РНК, была достигнута уже в работе [21]. Было показано, что при понижении концентрации нуклеозид-трифосфатов ниже 0,15mM прекращается синтез РНК (в отсутствие матрицы), хотя матрично-зависимый синтез РНК шел нормально. Кинетика синтеза в "безматричных" условиях имеет очень длинный лаг-период, в отличие от короткого при наличии матрицы. Однако, сомнения оставались. И только после серии блестящих исследований Биебричера, Эйгена и Льюс 1981-1987 годов [например, 4] "безматричный" синтез РНК был окончательно доказан.
И тем не менее, в некоторых исследованиях, где не смогли добиться требуемой чистоты экспериментов, пытаются выдать методические недостатки постановки исследований за их достоинства. К примеру, группа А.В.Четверина, найдя в продуктах "безматричного" синтеза РНК фрагменты, гомологичные 23S РНК E.coli и B.subtilis, а также гомологи фрагментам РНК из фага Qb, утверждает, что все эксперименты в этой области объясняются исключительно присутствием в реакционных смесях контаминирующих экзогенных РНК из воздуха лабораторий, как это имело место в их постановке. В качестве решающего аргумента демонстрируются чашки Петри с агарозой, содержащей Qb репликазную систему. Чашки выдерживают в открытом виде от 0 мин. и 10 мин. до 1 часа. Продукты реакции окрашивают бромистым этидием на РНК. После этого фиксируют нарастающее количество РНК, свидетельствующее о посеве загрязняющих посторонних РНК из лабораторного воздуха и их автокаталитическом размножении [17, 9]. То, что из грязного воздуха в лабораторную посуду может попасть все, что угодно, включая РНК, не удивительно. Но это никак не относится к исследованиям, которые проводятся на высоком методическом уровне. Удивительно другое. Биебричер и соавторы, получившие в течение 10 лет безукоризненные ключевые результаты по "безматричному" синтезу РНК в очищенных системах in vitro, не считают этот феномен нарушением центральной догмы молекулярной биологии и генетики. Они полагают, что в процессе энзиматической Qb-полимеризации рибонуклеозид-трифосфатов in vitro возникают некие наборы пробных РНК (низкомолекулярная фракция 6S РНК), некоторые из которых, являясь сами для себя матрицами, автокаталитически самореплицируются и при этом мутируют. Мутационные варианты подвергаются естественному отбору в духе теории Дарвина и после нескольких раундов размножения микроэволюция синтезируемых РНК прекращается на РНК-победителях. Вероятно, для Биербричера и многих других отказаться от центральной догмы означает полную смену, или существенное дополнение представлений о стратегии работы хромосом. Оснований для этого они пока не находят. Но и объяснить ими же полученные результаты они тоже не в состоянии. И прежде всего, сам факт синтеза РНК без ДНК. Они признают, что не понимают биологическую роль стабильно синтезируемой в таких системах фракции 6S РНК.
При биосинтезе in vivo, в процессе размножения фага Qb в E.coli, 6S РНК также образуется, она, как и in vitro, гетерогенна по последовательностям нуклеотидов и вариабельна по их числу: от 100 до 200. При каждом "безматричном" и нормальном синтезе образуются разные наборы РНК. И только некоторые из них реплицируются по (-) цепям. Следовательно, с самого начала есть отбор матриц на размножение, и следовательно, они не случайны по "текстам" (семантике). Биологическая роль 6S РНК не известна потому, что они не кодируют белки и не вовлечены в инфекционный процесс. Это ограничение в понимании функций нуклеиновых кислот продиктовано существующей жесткой парадигмой, что генетические структуры работают исключительно на вещественном уровне, что оспаривается современными данными. Существенная деталь - очень долгий лаг-период при синтезе 6S РНК, доходящий до 16 часов в случае ДНК-зависимой РНК- полимеразы у фагов Т7, Т3 и SP6 [Интернет]. Этому также объяснений нет. И еще один нетривиальный факт. Qb-репликаза состоит из 5 различных белковых субъединиц, названных I-IV или субъединицы ,,,. Пятая названа хозяйским фактором (host factor HF). Субъединица I идентифицирована как рибосомальный белок S1, а субъединицы III и IV являются факторами трансляции и элонгации EF-Tu и EF-Ts. Субъединицы I, III и IV работают при рибосомальном синтезе белков, но в данном случае используются фагом Qb для синтеза РНК. Существует мнение, что 6S РНК, размножаясь в бактериях как "молекулярный паразит", является "эгоистической РНК" [13]. Иными словами, 6S РНК можно считать аналогом "эгоистической ДНК" эукариот, роль которой также не совсем понятна. Версия функций "эгоистической ДНК" предложена, и она трактуется как один из способов вещественно-волновой кодировки пространственно-временной структуры организмов с биологическим использованием принципов голографии, солитоники, лингвистики, резонансно-волновых взаимодействий и лазерных процессов [26, 27]. Не исключено, что эти же механизмы в упрощенном варианте приложимы и к функциям 6S РНК бактерий. Кроме того, возможно, 6S РНК работает как своеобразная "антенная система", воспринимающая внешние физические поля как регуляторный генетико-волновой фактор (см. ниже).
ГИПОТЕЗА ВИРТУАЛЬНЫХ И ВОЛНОВЫХ ГЕНОВ
Таким образом, всем трем обсуждаемым явлениям - прионам, теломерам и Qb-репликазе - присуща общая стратегическая черта. Это их необычная способность реплицировать белки, ДНК и РНК, вроде бы, безматричным (безматериальным), и пока необъяснимым, путем. Для прионов - это непонятный путь проникновения из желудка в мозг и необъяснимая штаммоспецифичность без генома, для теломер - непонятный ALT механизм синтеза концевых хромосомных ДНК, а для Qb-репликазы - непонятный "безматричный" синтез РНК.
В качестве одного из объяснений мы выдвигаем гипотезу, что прионоподобные паразитические белки обладают виртуальным геномом, взятым “взаймы” у клетки-хозяина на момент размножения этих белков. Это размножение проходит не только и не столько за счет перехода PrPcPrPsc. Виртуальный геном может работать двумя путями (см. Рис.1 и 2).
Кето-группы белковых аминокислот PrPsc могут реагировать с ОН-группами остатков рибозы акцепторных концов соответствующих транспортных РНК (тРНК). Это каталитический процесс. В ходе соответствующих ферментативных реакций возникающий поли-тРНК-континуум попарно пространственно сближает антикодоны, образуя ковалентно дискретное “подобие информационной РНК” (пиРНК). Это этап почти обратный синтезу белка на рибосоме. И возможно, он проходит на А- , Р-сайтах рибосомы. Затем идет обратнотранскриптазный синтез ДНК на пиРНК. Для этого необходима соответствующая обратная транскриптаза, способная работать с ковалентно-дискретной матрицей пиРНК.
Возможен и другой путь (Рис.2), когда рестриктазами “срезается” антикодоновый политриплетный континуум тРНК с последующим энзиматическим “сшиванием” (лигированием) триплетов. Это также дает РНК-матрицу для синтеза ДНК. Таким образом создается клон молекул ДНК, которые могут реплицироваться (размножаться) или транскрибироваться в нормальные информационные РНК (иРНК), ответственные за синтез PrPsc.
В связи с этой гипотезой возникает вопрос о взаимоузнаваниях тРНК и белковых аминокислот, аналогичный вопросу, поставленному Вейсом и др. [23] о способности РНК аптамеров (РА) распознавать прионовые белки (см. выше). Ответа на него пока нет, но в контексте нашей версии прионового генома важнее другое - РА принципиально способны узнавать определенные аминокислотные последовательности и это косвенно подтверждает идею о способности тРНК узнавать белковые аминокислоты. А это необходимо для выстраивания линейного тРНК-континуума и всех последующих актов создания временного виртуального генома прионов.
В представленных вариантах геном, точнее часть белоксинтезирующего аппарата клетки-хозяина, “заимообразно” передается белкам PrPsc на время синтеза ДНК-РНК-матриц. По причине такого временного сосуществования PrPsc-РНК-ДНК комплексов, нуклеиновые кислоты не обнаруживаются в составе PrPsc в процессе препаративного выделении их в “чистом” виде. Заимообразность такого рода архаична, однако, при патологических состояниях биосистемы позволяет обойти энергетически и организационно тяжелый путь постоянного обращения к хромосомам для синтеза паразитических белков. Виртуальный геном PrPsc и аналогичных белков-паразитов придает им свойства вирусоподобности, штаммоспецифичности, которые зависят от особенностей полимеразных систем клетки-хозяина. Наверное, такой или такие механизмы размножения белков были эволюционными предшественниками вирусных инфекций и предвестниками воспроизведения первичных организмов. Этот палеобиохимический процесс клонирования белков, видимо, сохранен и может работать при многих патологиях (иммунодефицит, ревматизм и др.). Видимо, возможен и более сложный вариант считывание информации с ассоциатов белков, который предствален на Рис.3.
Если наша гипотеза подтвердится, необходимы будут дальнейшие поправки к догме молекулярной биологии об однонаправленности стратегического потока информации в биосистемах: ДНКРНКБЕЛОК. Первую поправку внесло открытие обратной транскриптазы. И схема стала иной: ДНКРНКБЕЛОК. Вероятно, следующим этапом будет следующая модификаци: ДНКРНКБЕЛОК. Нам представляется, что в случае паразитических белков-прионов рибосома, по сути, работает как белок-зависимая иРНК-полимераза. На Рис.4. дана обобщенная схема, модифицирующая центральную догму молекулярной биологии и генетики с учетом высказанных соображений.
Относительно проникновения PrPsc в мозг из желудка, минуя гемато-энцефалический барьер, можно полагать, что биосистемами используются вещественно-волновые механизмы памяти генома и дистантной солитонной и иной волновой трансляции геноинформации, которые предложены ранее [26, 27]. Косвенно в пользу вещественно-волновой памяти ДНК могут свидетельствовать данные о "безДНКовом" синтезе прионовых белков у дрожжей. Возможен также нейронно-волновой путь передачи информации о первичной структуре иРНК прионов высших биосистем через внутреннюю колебательную структуру солитонных пакетов нервных импульсов, идущих по волокнам от желудка в мозг. Этот способ свертки информации о РНК в спектр резонансов Ферми-Паста-Улама, промодулированных РНК-текстом, и перенос его спектра в структуру спайка потенциала действия в нервном проводнике недавно предложен [25].
К вещественно-волновым механизмам синтеза ДНК можно отнести и феномен ALT-удлинения теломер, и синтез de novo 6S РНК в Qb-репликазной "безматричной" системе.
Что из себя представляют вещественно-волновые информационные механизмы памяти (сокращенно "ВИМ") в генетических и других управляющих структурах организмов? Изучение их только начинается. Вероятно, некоторыми типами запоминания и формирования вещественно-волновых хромосомных образов-моделей, образов-программ являются голографическая и/или лингвистико-фоновая память генома [26, 27]. Надо полагать, что ВИМ-пути репликации информационных биополимеров разнообразны, имеют истоки, начиная с уровня элементарных частиц, и в первом приближении, их можно классифицировать следующим образом:
1. ВИМ электрона или "дырка". Это первый, классический пример из квантовой электродинамики, когда удаляемый объект (электрон) оставляет на месте своего пребывания определенную область, эквивалентную ему, но с противоположным, положительным, зарядом. Эту область Поль Дирак назвал "дыркой" [10]. Она ведет себя подобно протону, хотя и не тождественна ему. "Дырки" реализуют положительную проводимость, которая существенна в свойствах полупроводников при так называемых p-n переходах.
Ассоциативно-голографические ВИМ-отображения. Здесь образ объекта, который экспонировался в двух интерферирующих когерентных полях, может восстанавливаться в форме волновых фронтов при отсутствии самого объекта. Это явление изучено детально и широко используется в технике и искусстве.
Фантомный листовой эффект (ВИМ листа). Открыт В.Г.Адаменко в 1975 году методом газо-разрядной визуализации и воспроизведен во многих лабораториях мира, в том числе и в работе [26]. Эффект заключается в том, что при помещении живого листа растения в высоковольтное высокочастотное поле, которое имеет строго определенные параметры, редко, но достаточно воспроизводимо, удается зафиксировать светящееся (искровое) изображение целого листа, даже если у него отрезана некоторая часть (не более 10-15%). Механизм такого явления имеет квази-голографическую природу, присущую генетическому аппарату высших биосистем [26, 27]. Фантомные или ВИМ-боли, возникающие иногда у людей, утративших в результате травмы руки или ноги, связаны с голографической памятью коры головного мозга.
4. ВИМ-структура сигнала, распределенная в фоне (о Фоновом Принципе см. [27]). Например, информация о точном смысле (значении, сигнале) омонимов типа 'лук', 'рейд', 'коса' закладывается контекстом сообщения, в котором они фигурируют. Весь текст (контекст) выступает как фон, в котором, как в целом, находится точная информация о части, то есть о точной семантике омонима. Омонимическая неоднозначность в высшей степени характерна для так называемого генетического кода - фактически, кода белков. Одинаковые знаковые дублеты кодонов кодируют разные аминокислоты, третьи же нуклеотиды кодонов могут быть любыми и не коррелируют с кодируемой аминокислотой ("Вобл" концепция Ф.Крика). По этой причине перед рибосомой часто, практически всегда, возникает дилемма такого вида: как считывать третий нуклеотид кодона - так, чтобы получилось "белковое слово" 'суд' или так, чтобы получилось 'сук'. Мы полагаем, что такие высоко вероятные ошибочные (случайные вследствие омонимии знаковых дублетов кодонов иРНК) выборы "аминокислот-букв" на место в "белковом слове" корректируются рибосомным комплексом за счет фоново-контекстной, ассоциативной ориентации рибосомы, учитывающей всю последовательность иРНК или, иначе говоря, ее контекст (фон). Фоновый Принцип - чрезвычайно важное теоретическое положение для практической биологии и медицины и сельского хозяйства, в частности, для волновой генетики. Например, в медицине применение этого принципа приведет к принципиально новым лечебным методологиям работы с онкогенами и геномом ВИЧ: появляется реальная возможность понимания, почему эти знаковые генетические структуры начинают внезапно функционировать только в строго определенном контекстном нуклеотидном окружении, возникающем вследствии транспозиций онкогенов и генома ВИЧ и/или при изменении "метаболического контекста". Они омонимичны, то есть не "читаемы" и не "узнаваемы" для организма, как патогенные начала, вне определенного фонового нуклеотидного окружения. В этом смысле вся грандиозная, и непонятная доселе, проблема геномных транспозиций становится прозрачной как фактор многомерного проявления конкретных семантик для временно омонимичных и/или временно нулевых (типа псевдогенов) знаковых нуклеотидных последовательностей. Если Фоновый Принцип экстраполировать на ALT-механизм восстановления теломер, то можно считать утрату конца теломеры предельным случаем омонимии, известным в лингвистике и теории информации (теорема Шеннона). В соответствии с этой теоремой, когда часть слов, букв (сигналов) в тексте утрачена или искажена, они могут быть восстановлены, с учетом всего текста (контекста), в том числе и по фоново-ассоциативному принципу, который включает шенновский эффект как частный случай [32]. В каком-то смысле это аналогично восстановлению полного изображения объекта из части голограммы, поскольку образ объекта, точнее информация о нем, не локализована в какой-то одной части голограммы, но распределена по всему ее пространству. В ALT-механизме восстанавливаемый сигнал - это информация об утраченной последовательности нуклеотидов теломеры, а фон (контекст) является текстом ДНК хромосомы, граничащим с его утраченной частью. Даже если допустить, что в ALT работает принцип рекомбинационного удлинения теломер, то и он не может обойтись без "знания" о том, какой фрагмент ДНК надо вставить на место отсутствующего. А это "знание" так или иначе связано с фоново-ассоциативной памятью генома, что близко к голографической памяти хромосомного континуума, который мы постулировали ранее [26, 27].
5.ВИМ ДНК. Этот феномен обнаружен в 1985 году, в кратком варианте опубликован в 1991 [12] и более детально исследован в 1994 [26]. Аналогичные данные получены независимо группой американских исследователей под руководством Р.Пекоры в 1990 году [1]. Внешнее проявление ВИМ ДНК заключаются в том, что при использовании метода лазерной корреляционной спектроскопии, при изучении светорасеяния препаратами ДНК in vitro, обнаруживается необычный эффект. В светорассеяние вносит вклад некий фактор, не предусмотренный теорией светорассеяния. Природа феномена не ясна. Группа Р.Пекоры характеризует ВИМ ДНК как ".. mimicing the effect of dust" (MED-effect), то есть эффект, имитирующий пыль, хотя исследователями предприняты специальные меры, чтобы в препаратах ДНК не было посторонних частиц. Группа Р.Пекоры обнаружила этот феномен методом корреляционной лазерной спектроскопии, но не на тотальных высокополимерных фракциях ДНК, как в наших экспериментах, а на рестриктных фрагментах ДНК определенной длины. И в этом (рестриктном) случае ДНК тоже вела себя “аномальным” образом: зондирующие фотоны дифрагировали не только на полинуклеотидных цепях, но и на “посторонних частицах”, которых в препарате заведомо не было. Этот, никак не прокомментированный группой Р.Пекоры, эффект затруднил ей объяснение необычного характера светорассеяния ДНК с позиций, казалось бы, хорошо разработанной теории Цимма и Роуза для влияния динамики полимеров в водных растворах на зондирующий световой, в том числе и лазерный, пучок.
Близкие, но не тождественные, данные получили Матсумото и др. [15], когда прямым наблюдением зафиксировали эффект “аномального” броунирования флуоресцентно-меченых нативных молекул ДНК. Эта "аномалия" проявляется в том, что рассчитанные на основе теории Цимма-Роуза коэффициенты трансляционной диффузии для отрезков ДНК длиной 56значительно отличаются от визуально наблюдаемой и количественно оцениваемой диффузионной динамики ДНК. Вместе с тем, теория дает хорошее совпадение с экспериментами для других, например, абиогенных полимеров типа лавсана, полиэтилена и т.д.
Можно думать, что в экспериментах по лазерному зондированию ДНК, светорассеяние происходит по принципам ВИМ, то есть не только на вещественных реально существующих молекулах ДНК, но и на ее виртуальных волновых эквивалентах (следовых структурах), оставляемых броунирующими молекулами этого суперинформационного биополимера. Здесь уместна некоторая аналогия с голограммой, когда определенным образом зондируемый лазером материальный объект "записывается" в рассеянном им световом поле и создает свою волновую (виртуальную) копию, существующую уже независимо от объекта-прототипа. Не исключено, что эффекты ВИМ объясняются теорией физического вакуума [33], где фундаментально обосновывается идея генерации торсионных (аксионно-кластерных) эквивалентов физических тел. Напомним, что ВИМ-эффекты на уровне генома давно известны в скотоводстве (и не только) как Фактор Телегонии, когда наследуются признаки первого самца-родителя, при рождении потомства той же самкой от второго, третьего и т.д. отцов.
Что касается “аномалий” динамики ДНК, обнаруженных в работе [15], то в их варианте опытов может происходить акцепция внешних фоновых физических полей, влияющих на квазиспонтанную динамику ДНК. Этот возможный вклад не принимался в расчет авторами, поэтому обнаруженный аномальный эффект также оказался вне понимания. Механизм такого рода влияния фоновых физических полей на нелинейную динамику белков предложен ранее [28, 29]. Он основан на взаимодействии внешних физических полей с атомами металлов, входящих в состав активных центров многих белков. Аналогичные пути влияния внешних (фоновых) и внутренних (эндогенных для биосистем) физических полей на динамику ДНК, наверное, имеют место, поскольку сахаро-фосфатный остов ДНК содержит различные металлы, роль которых не установлена и может заключаться именно в волновых знаковых биоориентациях генома.
ВОЗМОЖНАЯ МОДИФИКАЦИЯ ЦЕНТРАЛЬНОЙ ДОГМЫ МОЛЕКУЛЯРНОЙ БИОЛОГИИ С ПОЗИЦИИ КВАНТОВОЙ НЕЛОКАЛЬНОСТИ ГЕНОМА И ЭКСПЕРИМЕТАЛЬНОЙ КВАНТОВОЙ ТЕЛЕПОРТАЦИИ
Тэги: гаряев, генетика
Природа феноменов ВИЧ и рака. Сложности в трактовке.
Сущность ВИЧ и рака лежит в той же области, что и сущность Жизни. Но мы до сих пор не понимаем главного в феномене Жизни – как она возникла на Земле и, каким образом она кодируется в хромосоомах. Существует ряд гипотез, каждая из которых, в лучшем случае, лишь часть правды. Отсюда теоретико-биологические трудности в трактовке ВИЧ и рака, а следовательно, и ошибки при их лечении. Оба болезнетворных начала оккупируют главное в любой биологической системе - ее генетический аппарат, то есть сумму “знаний” организма о самом себе. И вот парадокс, вроде бы о хромосомах и ДНК знаем достаточно. Онкогены обнаружены, геном ВИЧ изучен и как эти информационные структуры функционируют в хромосомах понятно. О работе генетического кода и рибосом, тоже вроде бы известно в деталях. Но этого почему-то оказывается недостаточно для разработки абсолютных методов борьбы с раком и ВИЧ.
Худшая ложь - это полуправда. В нее привычно верится, особенно, если это полуправда “знания” о генетическом кодировании. Здесь все - неприступный бастион для критики и все догматизировано. Даже основное понятие, стратегическая схема генетического кодирования (ДНКà РНКà Белок) так и названо "Центральная Догма". Атака на эту догму до недавнего времени казалась бессмысленной, обреченной на провал. И как оказалось, напрасно. От того правильно ли мы понимаем генетическое кодирование зависит правильность или неточность стратегии изучения ВИЧ и рака, да и не только этих патологий. Первую брешь в этой догме пробило открытие ревертазы, и догма автоматически превратилась лишь в версию, которая звучит теперь существенно скромнее: ДНКÛ РНКà Белок. Но и эта модификация наших представлений о биосинтезе белков подвергается эрозии, поскольку она - лишь очередное приближение к истине, к пониманию языково-образного плюрализма генома как средству кодирования пространственно-временной структуры биосистем [32, 33].
Что мы хотим доказать?
В данном исследовании мы развиваем свои идеи, цель которых не в окончательном разрушении так называемой “канонической” триплетной модели генетического кода, но в развитии и установлении ее точного места в системе знаний о принципах работы хромосом. Да, можно сказать, что триплетный код - истина. Но это истина такого рода, как если бы утверждали, что с помощью алфавита можно написать любое слово. Это верно. Но если мы пытаемся идти дальше, располагая только этим знанием, и доказываем, что с помощью алфавита можно конструировать правильные предложения, то это неверно. Потому, что для построения человеческой речи требуются законы мышления, грамматики и логики. А геном именно речеподобен и логичен, хотя эти фундаментальные черты - не единственный способ выражения его образно-смысловых конструкций. Более того, мы склонны принять идеи В.В.Налимова [43], которые приводят нас к идее, что геном обладает способностью к квазисознанию. Наши логика и модели – это попытка получить знания более высокого уровня о законах построения генетических текстов и иных знаковых структур генома, знания, которые находятся в начальной стадии развития. Основы их были заложены еще в 20-е годы русскими исследователями А.Г.Гурвичем [38], В.Н.Беклемишевым [29] и А.А.Любищевым [41].
Что же можно предложить в развитие и дополнение общепринятой теории генетического кодирования, и как это может помочь решить, как частный случай, проблему ВИЧ и рака? Условно, до полного доказательства, примем три положения, имеющие определенные теоретико-экспериментальные доказательства [8, 32, 33, 37].
Молекулы ДНК в составе хромосом обладают вещественно-волновой двойственностью, родственной дуализму элементарных частиц. В соответствии с этим ДНК кодирует организм двояко - и с помощью вещества ДНК, и за счет ее знаковых волновых функций, в том числе и на уровне собственных лазерных излучений [28].
Генетический аппарат обладает способностью быть нелокальным на молекулярном уровне (голографическая память хромосомного континуума) и квантово-нелокальным в соответствии с эффектом Эйнштейна, Подольского, Розена [4]. Последнее означает, что генетическая и иная регуляторная волновая информация генома записывется на уровне поляризаций его фотонов и нелокально (повсеместно и за нулевое время) передается (разворачивается) по всему пространству биосистемы по кодовому параметру поляризаций. Этим достигается безинерционный информационный контакт между миллиардами клеток организма.
Геном в целом и отдельные клеточные ядра обладают квази-сознанием разного уровня; и геном, и отдельные клеточные ядра способны генерировать и распознавать текстово-образные регуляторные структуры с использованием фонового принципа, голографии и квантовой нелокальности.
Что делать дальше?
Допустим, получены окончательные доказательства этих положений. Тогда проблема ВИЧ и рака представляется в совершенно ином интеллектуальном измерении. Попытаемся пояснить эти идеи и факты в теоретико-биологическом и медицинском осмыслении. Например, что такое вещественно-волновой дуализм ДНК и как он связан с многочисленными кодовыми функциями хромосом, существенно отличающимися от известного триплетного генетического кода? В определенном смысле геном выступает как сложный многоволновой лазер с перестраиваемыми частотами. Он излучает свет, гено-знаково промодулированный ДНК по амплитуде, фазе, частоте и поляризации. Более того, геном, вероятно, еще и разер, конвертирующий когерентные знаково поляризованные фотоны в телепортационно связанные с ними когерентные изоморфно знаково поляризованные радиоволны широкого спектра [37] (Гаряев П.П., Тертышный Г.Г., Леонова Е.А. и др. Радиоволновая спектроскопия локализованных фотонов: выход на квантово-нелокальные биоинформационные процессы. Датчики и системы (N9 стр.2-13 за 2000 год).). Геном является также и мобильной меняющейся мультиплексной квазиголограммой, способной при ее многоволновом автосчитывании собственными фотонными излучениями образовывать свето-радиоволновые генознаковые и иные регуляторные структуры [37]. Такие структуры являются реестрами электромагнитных разметочных схем (калибровочных полей) построения пространства-времени биосистем. И наконец, геном – это квазитекстовое образование, с элементами квантовой нелокальности, которое может само себя безинерционно "прочитывать" в миллиардах своих клеток и пользуется полученной при этом информацией как одним из руководств жизнедеятельности и одним из способов организации своей структуры [8, 37]. Наверное, эти идеи о новых информационных измерениях генома для многих биологов и генетиков, а тем более для медиков, воспринимаются сейчас как "китайская грамота". Однако, не для всех из них. Мышление такого рода, пустившее корни в 20 годах в России, набирает силу с резким ускорением в последнее десятилетие.
Из этого с определенностью следует, что пока не поздно, необходимо менять стратегию поиска методов лечения ВИЧ и рака, поскольку традиционные попытки решить эти проблемы все более похожи на желание снять богатый урожай, засевая асфальт. Новая стратегия должна быть основана на фундаментальных исследованиях вещественно-волновых и квази-речевых атрибутов генома высших биосистем. Еще раз подчеркнем, что мы рассматриваем хромосомный континуум как знаковый лазерно-радиоволновой излучатель [8, 33, 37]. И этому есть прямые экспериментальные свидетельства. Например, для доказательства лазерных потенций генетических структур мы показали, что in vitro ДНК и хроматин может быть накачан как лазеро-активная среда с последующей лазерной генерацией света [28].
Если мы знаем эти существенные характеристики генома, то возникают естественные и конкретные вопросы – меняется ли знаковый характер лазерно-радиоволновой излучений хромосом при встраивании в них чужеродного генома ВИЧ? И что при этом происходит с "семантикой" излучений при транспозициях онкогенов или любых других мобильных полинуклетидных последовательностей, а такжже при B« Z и иных конформационных переходах в ДНК in vivo? Связано ли это с изменением квази- и голографических программ, то есть, создаются ли новые, изменяются ли старые, “стираются” ли они, и так далее? Динамичен ли при таких перестройках генома параметр поляризации излучений в семиотическом аспекте? Отражается ли все это на работе рибосом? Такие вопросы можно множить и далее. Ответ на любой из них может стать ключевым в трактовке природы ВИЧ и рака.
Более подробно о теоретичеких построениях.
Или другая проблема, также фундаментальная. Онкогены и геном ВИЧ, занимая определенные позиции в трехмерном пространстве хромосом клеток-хозяев, до какого-то времени не проявляют себя как патогенные факторы. В этом смысле поведение ВИЧ в организме инфицированного человека непредсказуемо. Инкубационный период ВИЧ может варьировать от недели до 10 лет. Считается, что есть определенный механизм индуцирования ВИЧ инфекции из латентного "спящего" состояния, но этот механизм не понят, а следовательно, упускается возможность заставить ВИЧ быть "спящим" в организме человека всегда. Организм и клетки их "не замечают" или даже, как в случае онкогенов, используют в благих целях как факторы роста. Почему они воспринимаемы адекватно (правильно) организмом до времени "X" и семантически перерождаются, вызывая управленческую катастрофу в клетках, после времени "X"? Следуя нашей логике, можно думать, что в патологических состояниях, равно как и в норме, работают, как минимум, четыре фактора - "голографичность" и "лингвистичность" генома, его фоновая (контектная) самоорганизация и его квантовая нелокальность.
Эволюция биосистем создала их собственные генетические "тексты" и геном-биокомпьютер как квази-разумный "субъект". Он на своем уровне "читает и понимает" эти тексты. Чрезвычайно важно для обоснования этой элементарной "разумности" генома, что естественные (не существенно на каком языке) человеческие тексты и генетические "тексты" имеют сходные математико-лингвистические и энтропийно-статистические характеристики. Это относится, в частности, к такому понятию как фрактальность распределения плотности частот встречаемости букв (для генетических "текстов" буквы – это нуклеотиды) [21].
Еще одно подтверждение лингвистической трактовки кодовых функций генома получено американскими исследователями [20]. Работая с “кодирующими” и “некодирующими” последовательностями ДНК эукариот (в рамках старых представлений о генах), они пришли к выводу, сходному с нашим и противоречащему догме о том, что знаковые функции сосредоточены только в белок-шифрующих участках ДНК. Они применили метод статистического анализа естественных и музыкальных текстов, известный как закон Ципфа-Мандельброта, а также известный постулат избыточности текстовой информации Шеннона, рассчитываемый как энтропия текстов (относительно энтропии текстов и статистики распределения слов в текстах см., например, [1, 25, 27, 31]. В результате они получили, что “некодирующие” районы ДНК (спейсовые, интронные и другие) более схожи с естественными языками, чем “кодирующие”. Исходя из этого, авторы предполагают, что “некодирующие” последовательности генетических молекул являются основой для одного (или более) биологических языков. Кроме того, авторами был разработан статистический алгоритм поиска кодирующих последовательностей ДНК, который выявил, что белок-кодирующие участки обладают существенно меньшими дальнодействующими корреляциями по сравнению с зонами, разделяющими эти участки. Распределение ДНК-последовательностей оказалось настолько сложным, что использованные методы переставали удовлетворительно работать уже на длинах, превышающих 103 - 102 пар оснований. Распределение Ципфа-Мандельброта для частот встречаемости “слов” с числом нуклеотидов от 3 до 8 показало большее соответствие естественному языку некодирующих последовательностей по сравнению с кодирующими. Напомним, что кодирование авторы понимают как запись информации об аминокислотной последовательности, и только. И в этом парадокс, заставивший их заявить, что некодирующие регионы ДНК - это не просто “junk” (в переводе с английского - “мусор”), а языковые структуры, предназначенные для каких-то целей с неясным пока назначением. Дальнодействующие корреляции в этих структурах авторам также не понятны, хотя и обнаружена нарастающая сложность некодирующих последовательностей в эволюции биосистем. И это продемонстрировано на примере семейства генов тяжелой цепи миозина при переходе от эволюционно низких таксонов к высоким. Приведенные данные [20] полностью соответствует нашим идеям, высказанным ранее и независимо [32, 33], о том, что “некодирующие” последовательности ДНК, а это около 95 - 98 % генома, являются стратегическим информационным содержанием хромосом. Оно имеет материально-волновую природу и поэтому многомерно и выступает, в сущности, как голографическая ассоциативно-образная и одновременно как семантико-семиотическая программа эмбриологического начала, смыслового продолжения и логического конца любой биосистемы. Интуитивно поняв тупиковость старой модели генетического кодирования, авторы [20] с ностальгической грустью прощаются со старой и хорошо послужившей моделью генетического кода, не предлагая, правда, ничего взамен.
Омонимо-синонимические неоднозначности генетических текстов. Для чего они нужны организму?
Общим фундаментальным семиотико-семантическим свойством естественных и генетических текстов является их синонимии и омонимии. Это обеспечивает для хромосом, также как и для естественных текстов и речи, сверхизбыточность информации, ее многозначность, а поэтому приспособительную гибкость. Многозначность одних и тех же генетических текстов переходит в однозначность за счет эффекта меняющегося положения ДНК-последовательностей в пространстве генома посредством их транспозиций и/или транспозиций их окружения. А это аналог ситуаций в естественных текстах и речи, когда синонимо-омонимические неоднозначности частей семантического поля снимаются контекстом (фоном; фоновый притнцип см. в [44]). В традиционной триплетной модели генетического кода легко обнаруживается омонимии кодирующих дублетов. Значение таких омонимий до сих пор не понято и не оценено, за некоторым исключением [33, 35]. Необъяснимая проблема омонимий кодонов информационных РНК (mРНК) сразу возникла при создании триплетной модели шифровки аминокислот в процессах биосинтеза белков. И сразу стала миной замедленного действия, поскольку правильное объяснение биологического (информационного) смысла таких омонимий автоматически приводит к необходимости существенного уточнения триплетной модели, если не сказать полного ее пересмотра. В чем проявляются омонимии кодонов? Ряд различающихся аминокислот кодируются одинаковыми дублетами в составе кодонов mРНК, а третьи нуклеотиды в кодонах могут меняться хаотически, они "воблируют" (англ. "wobble" – качание) и могут быть любыми из 4-х канонических. Вследствие этого они не коррелируют с кодируемыми аминокислотами [3, 11]. По этой причине возникает семантическая неоднозначность выбора рибосомой антикодонов транспортных РНК (tРНК), несущих аминокислоты. Например, каждый из синонимических кодонов стандартного кода высших биосистем AGT и AGC кодирует серин, а каждый из синонимических кодонов AGA и AGG кодирует аргинин. Поскольку третьи нуклеотиды кодонов mРНК в сочетании со знаковым дублетом не имеют точных аминокислотных коррелятов, а первые два знаковых нуклеотида кодонов одинаковы, но при этом кодируют разные аминокислоты, возникает неоднозначность в выборе антикодонов tРНК. Иными словами, рибосома с одинаковой вероятностью может выбрать и сериновую, и аргининовую tРНК, что может привести к синтезу аномальных белков. В действительности же таких ошибок не происходит, точность белкового биосинтеза чрезвычайно высока. Только в некоторых метаболически аномальных ситуациях происходят ошибки (наличие некоторых антибиотиков, аминокислотный дефицит и др.). В норме рибосома каким-то образом делает правильный выбор антикодонов tРНК из омонимичных дублетов.
Мы полагаем, что правильный выбор из дуплетных антикодонов-омонимов реализуется по резонансно-волновым и контекстным (ассоциативным, голографическим) и/или так называемым “фоновым механизмам”. Омонимичность аминокислотного кода может быть преодолена точно так же, как это происходит в естественных языках - путем помещения омонима, как части, в целое, то есть в законченную фразу, контекст которой дешифрует омоним и присваивает ему единственное значение, создавая однозначность. Поэтому mРНК в качестве своего рода “фразы” или “предложения” должна работать в белковом синтезе как функциональное кодирующее целое (нелокально), задающее последовательность аминокислот на уровне ассоциатов аминоацилированных tРНК, которые комплементарно взаимодействуют со всей молекулой mРНК. Макростерические несоответствия между mРНК- и tРНК- континуумами могут сниматься за счет конформационной лабильности макромолекул. При этом роль А-Р участков рибосомы заключается в акцепции таких ассоциатов - предшественников белка с последующей энзиматической сшивкой аминокислот в пептидную цепь. В этом случае будет происходить контекстно-ориентированный однозначный выбор и снятие омонимии дуплет-антикодонов. Можно предсказать в связи с этим, что взаимодействие аминоацилированных-tРНК с mРНК носит коллективный фазовый характер по типу реассоциации (“отжига”) однотяжных ДНК при понижении температуры после “плавления” нативного полинуклеотида. Существуют ли экспериментальные данные, которые можно было бы трактовать в таком духе? Их немало и они сведены в обзорно-аналитическом исследовании [45]. Приведем некоторые из них. Известно, что правильность узнавания молекулами tРНК терминирующих кодонов зависит от их контекстного окружения (подтверждение наших теоретических построений), в частности, от наличия за стоповым кодоном уридина. Так, в работе [9] показано следующее. Вставка строки из девяти редко используемых CUA-лейциновых кодонов после 13-го в составе 313 кодонов тестируемой mРНК сильно ингибируют их трансляцию без явного влияния на трансляцию других mРНК, содержащих CUA-кодоны. Здесь явно проявляются контекстные ориентации трансляции. Хорошо просматривается стратегическая линия влияния строго определенных и далеко расположенных от места образования пептидной связи кодоновых вставок в mРНК на включение или не включение определенной аминокислоты в состав синтезируемого белка. Это дистантное влияние, связанное с континуальностью белкового синтеза (также пример нелокальности функций генетического аппарата), когда mРНК воспринимается белок-синтезирующим аппаратом не только по частям (понуклеотидно, локально), но и как целое (нелокально). Однако, в в цитируемой работе этот ключевой феномен просто констатируется, оставаясь для исследователей непонятным и, видимо поэтому, даже не обсуждается. Таких работ становится все больше. В той, что мы обсуждаем, ссылаются, к примеру, на полдюжины аналогичных результатов, где трактовка в этом смысле также затруднена. Причиной этому, понятно, является несовершенство модели триплетного генетического кода. Модель не точна еще и потому, что существуют необычные протяженные (swollen) антикодоны. Когда они вовлечены в синтез белка, в А-сайте рибосомы участвуют не три, а большее количество пар оснований [45]. Это означает, что догматический постулат триплетности кода нарушается и в этом случае. В [45] приводятся результаты исследований по взаимодействию tРНК-tРНК на рибосоме, и это также полностью подтверждает нашу идею об ассоциате (континууме) нагруженных аминокислотами tРНК как предшественнике белка. В [45] высказана существенная мысль, близкая нам, что эффект действия контекста mРНК на однозначное включение аминокислот в пептид является отражением неких фундаментальных, и пока плохо изученных, закономерностей декодирования генетической информации в процессе белкового синтеза. Напомним, что генетическая информация о синтезе белков занимает всего около 1% объема хромосом. 99% заняты программами существенно более высокого уровня.
Прионы – последний удар по центральной догме молекулярной биологии.
Как видим, ранние представления о генетическом коде и знаковой работе белок-синтезирующего аппарата носят упрощенный характер. Возможно, последним аргументом в пользу окончательного пересмотра центральной догмы молекулярной биологии служит феномен прионов. Прионы – это низкомолекулярные паразитические белки (PrPsc), поражающие головной мозг животных (бешенство коров) и человека (болезнь Альтсгеймера, синдром Крейцфельда-Якоба и другие). Необъяснимой особенностью прионов является их вирусоподобная штаммоспецифичность. Но штаммоспецифичность присуща только микроорганизмам или вирусам, которые располагают генетическим аппаратом. Вместе с тем считается, что геном у прионов отсутствует, поскольку все попытки обнаружить в составе прионов хотя бы следы ДНК или РНК заканчивались неудачей. Возникает сильное противоречие, которое в очередной раз ставит под сомнение центральную догму молекулярной биологии: генома у прионов нет, а генетические признаки налицо. Не в силах объяснить это, “спасая” центральную догму, все-таки предполагают, что остатки ДНК или РНК прячутся в каких-то складках молекул прионов [10]. Однако, десятилетия исследований прионов, увенчавшиеся нобелевской премией 1997 года Стенли Прузинеру за исследования в этой области, показали абсолютно точное отсутствие в их составе нуклеиновых кислот, а следовательно и генома [23]. Как выйти из этого противоречия? Если придерживаться центральной догмы, то это невозможно. Отказавшись от нее, мы можем представить себе следующий сценарий биогенеза прионов [34]. Главной знаковой фигурой здесь выступает "виртуальный геном прионов", то есть геном временный, взятый заимообразно у клеток-хозяев. Точнее можно сказать, что это белок-синтезирующий аппарат клеток-хозяев. Похоже, прионы в качестве одного из способов размножения сохранили палеогенетический путь, позволяющий им в ряде случаев не пользоваться генами, кодирующими их в хромосомах, а саморазмножаться иным путем, игнорируя центральную догму молекулярной биологии и генетики. Для клетки, чтобы синтезировать прионы, обращаться к их генам - это, хотя и прогрессивный, но организационно и энергетически трудный путь. Прионы могут поступать проще. Мы предполагаем, что NH-группы пептидных связей PrPsc могут реагировать с ОН-группами остатков рибозы акцепторных CCA-последовательностей соответствующих tРНК. В ходе гипотетических ферментативных реакций возникающий поли-tРНК-континуум, колинеарный PrPsc, попарно пространственно сближает антикодоны, образуя ковалентно дискретное “подобие информационной РНК” (пиРНК). Это этап почти обратный синтезу белка на рибосоме. Вероятно, он проходит на А- , Р-сайтах рибосомы. Затем идет синтез РНК на пиРНК. Для этого необходима соответствующая РНК-полимераза, способная работать с ковалентно-дискретной матрицей пиРНК. В этом и заключается "заимообразность" пользования белок-синтезирующим аппаратом клеток-хозяев на время размножения прионов. Такая временность создает иллюзию отсутствия у них генетического аппарата. При этом пептидные цепи прионов служат матрицами, на которых попарно на А-Р участках рибосом, выстраивается поли-tРНК-континуум, образуя дискретные полиантикодоны. Последние, объединяясь попарно, либо сразу служат матрицей для РНК-зависимого синтеза mРНК прионов, либо (в другом варианте) полиантикодоны за счет специфического сплайсинга вырезаются с последующим лигированием в ковалентно непрерывную матрицу mРНК прионов. Далее mРНК прионов на рибосоме полимеризует сами прионы. Это означает, что рибосома работает в обратном направлении и является при этом "прион-полиантикодон-зависимой mРНК полимеразой". И соответтсвенно, в нарушении догмы, информация идет от белка к РНК. Тогда схема догмы окончательно видоизменяется: ДНКÛ РНКÛ Белок. Тогда это уже не догма, а просто рабочая схема, с которой надо работать, уточняя и развивая ее. Штаммоспецифичность прионов, при таком взгляде на их биогенез, объясняется особенностями обратной работы рибосом, временно рекрутируемых при синтезе каждого из штаммов прионов. А эти особенности определяются таксономическим положением биосистем, продуцирующих прионы.
Вернемся вновь к пока еще общепринятым основным положениям модели генетического кода: он является триплетным, неперекрывающимся, вырожденным, не имеет “запятых”, т.е. кодоны ничем не отделены друг от друга. Поток информации идет от ДНК к РНК и далее к белку. И наконец, код универсален. Что осталось от этих положений? Фактически ничего. В самом деле, код, видимо, является двух-, трех-, четырех- ... n-буквенным как фрактальное и гетеромультиплетное образование, кодирующее не только отдельные белки, но и функционально связанные ассоциаты белков. Он перекрывающийся за счет сдвигов рамок считывания рибосом. Он имеет запятые, поскольку гетерокодоны могут быть отделены друг от друга последовательностями с иными функциями, в том числе с функциями пунктуации. Код не универсален - в 14 случаях он отличен от стандартного кода высших биосистем. Эти случаи относятся к митохондриальному, дрожжевому, микоплазменному, трематодному и другим кодам низших организмов [5, 6].
И последнее: белок может служить матрицей для РНК, как мы видим на примере прионов. Как понимать реальный генетический, а точнее белковый, код с учетом приведенных противоречий и предлагаемой нами логики рассуждений? Можно постулировать качественную, упрощенную, первичную версию вещественно-волнового контроля за порядком выстраивания аминокислот, задаваемым ассоциатами аминоацилированных tРНК как предшественниками белков. Приняв такую версию, легче понять работу белкового кода как одной из множества иерархических программ вещественно-волновой организации биосистемы. В этом смысле такой код - первый этап хромосомных планов построения биосистемы, поскольку язык генома многомерен, плюралистичен и не исчерпывается задачей синтеза протеинов. Основные положения предлагаемой ориентировочной модели вещественно-волновых знаковых процессов при биосинтезе белков сводятся к следующему:
Многокомпонентный рибонуклеопротеидный белоксинтезирующий аппарат является системой генерации высокоорганизованных знаковых излучений акустико-электромагнитных полей, стратегически регулирующих его самоорганизацию и порядок включения аминокислот в полипептидную цепь.
Аминоацилированные tРНК ассоциируют в последовательности - предшественники синтезируемых белков до контакта с А-P участком рибосомы. При этом континуум антикодонов пулов tРНК комплементарен всей mРНК, за исключением дислокаций, определяемых наличием не канонических нуклеотидных пар.
Порядок чередования аминоацилированных tРНК в ассоциатах-предшественниках белков определяется знаковыми коллективными резонансами всех участников синтеза аминокислотных последовательностей. Ключевые волновые матрицы здесь пре-mРНК и mРНК, работающие как целостный континуум (макроконтекст) разномасштабных по длине гетерополикодонов, включая интронную фракцию пре-mРНК. Главная функция волновых матриц - ассоциативно-контекстная ориентация последовательности аминоацилированных tРНК, ориентация, в большей степени чем "вобл-гипотеза" Ф.Крика, игнорирующая правила канонических спариваний нуклеотидов в одномерном пространстве mРНК-tРНК. На рибосоме, в дополнение и/или наряду с резонансными регуляциями взаимного расположения кодон-антикодоновых континуумов функционируют лазероподобные излучения участников данного процесса, корригирующие порядок включения аминокислотных остатков в пептид. Рибосома энзиматически ковалентно фиксирует “де-юрэ” пептидные связи аминокислотных последовательностей, намеченные “де-факто” в полиаминокислотном-поли-tРНК-ассоциате, как предшественнике белка.
Резонансно-волновая “цензура” порядка включения аминокислот в пептидную цепь устраняет потенциальный семантический произвол создания ошибочных белковых “предложений”, следующий из омонимии семейств кодонов, и обеспечивает их правильное “аминокислотное осмысление” за счет контекстного снятия омонимии неоднозначных одинаковых дублетов в кодонах. Тот же механизм работает при неоднозначностях более высокого порядка, когда число кодонов (n+1).
Вырожденность генетического кода необходима для пре-mРНК-mРНК-зависимого контекстно-ориентированного точного подбора аминоацилированных tРНК, определяемого характером волновых ассоциативных резонансных взаимодействий в белок-синтезирующем аппарате.
Один из механизмов процесса создания безошибочных последовательностей аминоацилированных tРНК на волновых матрицах пре-mРНК-mРНК можно рассматривать как частный случай частично комплементарной реассоциации однотяжных ДНК-ДНК и РНК-ДНК или, в более общем случае, как акт самосборки, известный для рибосом, хромосом, мембран и других молекулярно-надмолекулярных клеточных структур.
Рибосома способна работать в направлении синтеза РНК на матрице белков.
Таким образом, роль mРНК знаково многовекторна и дуалистична. Эта молекула, как и ДНК, в эволюции знаменует собой узловое событие - взаимодополняющее синергичное единство вещественной и волновой геноинформации. Неоднозначность вещественного кодирования снимается прецезионностью волнового, которое реализуется, вероятно, по механизмам коллективных резонансов и лазерно-голографических (ассоциативных, контекстно-фоновых) эффектов в клеточно-тканевом континууме. Скачок к более развитому волновому регулированию трансляции РНК® Белоксопровождается частичным или полным отказом от правила канонического спаривания аденина с урацилом (тимином) и гуанина с цитозином, свойственного эволюционно ранним и более простым этапам репликации ДНК и транскрипции РНК. Такой отказ информационно необходим, неизбежен и энергетически предпочтителен на уровне высших биосистем. Еще раз подчеркнем, что контекстные ассоциативно-голографические механизмы работы белок-синтезирующей системы организмов теснейшим образом связаны с так называемым “фоновым принципом” [44], а также, вероятно, с многовекторной и многосмысловой логикой управления сложными системами (кенограмматика Герхарда Томаса) [26]. С этой позиции макроконтексты пре-информационных и контексты информационных РНК можно рассматривать как фон, который в данной ситуации и в данной трактовке является "шумовым источником информации". Это обеспечивает резкое усиление сигнала, по которому происходит точный выбор (волновое распознавание) одной из двух омонимичных аминоацилированных tРНК, одна и только одна из которых должна войти в точные белковые “фразу” или “слово”. Этот выбор возможен после выделения когерентной составляющей в форме повторов одних и тех же "осмыслений" (распознаваний) рибосомой одного из двух одинаковых дублетов в кодонах. Ситуацию можно пояснить на простом примере. Скажем, в предложении надо выбрать одно из двух слов (аналогов кодонов с дублетами-омонимами). Эти слова - “суд” и “сук”. Ясно, что выбор зависит от целого предложения, от контекста, который выступает как фон, позволяющий выделить сигнал - нужное слово. Если предложение звучит “я увидел толстый сук на дереве”, то замена здесь слова “сук” на “суд” будет равносильна введению шума и потере сигнала. Вероятно, аналогична роль пре-информационных РНК и интронов - это различные уровни контекстов, которые должны быть каким-то образом “прочитаны” и “осмыслены” живой клеткой и ее рибосомным аппаратом для принятия точного решения по выбору антикодона tРНК в ситуации омонимии.
Аппаратом континуального (нелокального) “чтения” контекстных РНК-последовательностей как целого может выступать многоликое семейство солитонов - оптических, акустических, конформационных, вращательно-колебательных и иных, возбуждаемых в полинуклеотиде. Функции таких солитонов могут выступать как способы накопления семантической информации о РНК-контекстах и следующей за этим смысловых регуляций кодон-антикодоновых знаковых взаимоотношений. Смысловые оценки при этом проводятся геномами-биокомпьютерами клеток. В качестве одного из способов континуального прочтения полинуклеотидов можно представить именно солитонный, сканирующий последовательность РНК. Например солитоны бегущих крутильных колебаний нуклеотидов на сахаро-фосфатной оси, физико-математически рассмотренный нами для однотяжных РНК-подобных участков ДНК [30, 36]. Такие солитоны реагируют на изменения последовательностей нуклеотидов модуляциями своей динамики, которая приобретает черты знаковости и может, вероятно, передаваться дистантно, то есть на расстояния, существенно превышающие длину водородных связей. Без дальней (волновой, континуальной) миграции сигнала о целом, то есть о пре-mРНК-mРНК-последовательностях, невозможна реализация ассоциативно-контекстных регуляций синтеза белков. Для этого необходима волновая способность солитонов (а также и голографической памяти) работать не только с частями, но и с целым. Такая континуальность, или что одно и то же, нелокальность, обеспечивает узнавание и правильный выбор рибосомным аппаратом истинного кодона из двух дублет-омонимичных, кодона псевдо-зашумленного фоном (контекстом).
Проведенные исследования, казалось бы, не связаны. Однако это не так. Процесс образования фантомов ДНК был обнаружен нами еще в 1984 г. Обнаружение и регистрация волновых реплик ДНК in vitro подтвердила эти результаты. Это еще раз подтверждает нашу гипотезу о способности ДНК быть структурой волнового автореплицирования и автосканирования внутриклеточного метаболического статуса, а в итоге всего организма с целью биокомпьютерного регулирования жизнедеятельности. Это полностью соответствует основному положению нашей концепции «волнового генома»: хромосомный континуум работает как стратегическая знаковая система на 2-х уровнях — вещества и поля одновременно, реализуя свои функции в качестве квазиразумной системы.
Однако это не единственная волновая система саморегуляции многоклеточных организмов. Вторая система основывается на когерентных излучениях хромосом в Уф-, видимой и ИК- областях спектра. Это дает возможность хромосомному аппарату использовать принципы голографической свертки-развертки генетической информации, квантовую нелокальность ее, а также лингвистические особенности работы дуэта «рибосома-иРНК» в рамках понятия квазиразумности генома как биокомпьютера..
Сейчас сложилась парадоксальная и многообещающая ситуация в генетике, молекулярной биологии и, соответственно, в медицине. Изучен геном человека. На это понадобились 10 лет усилий программы «Геном человека», и теперь известна последовательность всех букв в нашем генетическом тексте.
Трансгенная инженерия набирает силу, и уже множество растений, животных и бактерий являются носителями искусственно внедренных в них полезных для человека генов. Сделаны первые шаги в клонировании животных с перспективой получить первый клон человека.
Но вот парадокс — чем выше наши технологические успехи в генетике и молекулярной биологии, тем дальше мы от всеобъемлющего понимания принципов работы генетического аппарата. Все несомненные успехи в этой области преимущественно связаны с функциями генов, которые кодируют белки. Эти гены занимают около 2% всей генетической памяти хромосом. Остальная, главная часть, т.е. 98%, до сих пор является непонятой генетиками и, вероятно, поэтому они назвали ее «мусором». Есть гипотезы, но все они призваны оправдать существование «мусорной» ДНК как некоего помощника для 2% «кодирующей» ДНК. Либо «мусор» трактуют как «кладбище вирусов»(!)
Игнорировать или так наивно понимать роль 98% генома — явная ошибка. Более того, правильно ли мы видим истинную роль 2% генетической информации, если 98% её для нас фактически — «terra incognita»? Понимаем слабо, поскольку не можем полностью излечивать рак, не можем противостоять ВИЧ, не можем бороться против туберкулеза, не можем продлить жизнь людей, по крайней мере, до рубежа 200 лет и т.д. Обещания генетиков обернулись опасными для жизни трансгенными продуктами питания, грозящим дисбалансом биосферы.
Клонирование животных дает нам, в основном, тщательно скрываемых уродов, или животных, которые аномально быстро стареют и умирают, как знаменитая овечка Долли.
Вполне естественно, что научное сообщество, например, в лице большой группы шведских ученых, начинает бить тревогу по этому поводу http://www.psrast.org/defknthe.htm. Где выход из этого странного состояния обилия экспериментального и противоречивого материала и дефицита достаточно полного теоретического осмысления принципов работы хромосом? Та же группа шведских ученых полагает, что одним из прорывных научных направлений являются наши разработки http://www.rialian.com/rnboyd/dna-wave.doc.
Суть наших идей, и что они дают практике, сводится к следующему. Мы исходим из очень простых стратегических соображений. Для успеха в попытках кардинально лечить людей и резко затормозить процессы их старения надо понять языки, на которых клетки общаются друг с другом. В какой-то степени нам удалось это сделать. Оказалось, что это языки тех самых 98% «мусора» в нашем собственном генетическом аппарате http://www.self-managing.net/genetica/Zip/V_Genetica%20(Kniga).zip.
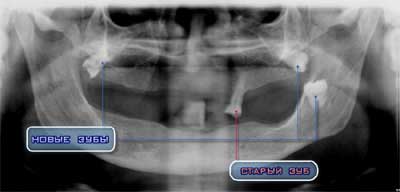
Уникальный рентгеновский снимок ротовой полости пациентки 60 лет – зубы, выросшие с наклоном под постоянно носимым протезом
Основным из них является язык голографических образов http://www.self-managing.net/genetica/Zip/Nestazionarn_golografia.zip, основанный на принципах лазерных излучений генетического аппарата http://www.self-managing.net/genetica/Zip/DNK%20laser.zip, работающего как квазиразумная структура http://www.rialian.com/rnboyd/dna-wave.doc Важно, что нашему генетическому аппарату свойственны реальные процессы, существенно дополняющие триплетную модель генетического кода http://www.self-managing.net/genetica/Zip/Central%20Dogma.zip. В этих процессах используются ранее неизвестные для организмов свойства http://www.self-managing.net/genetica/Zip/FROL.zip и др.
Что нам дают эти новые знания? Ответ таков: теперь, зная об этом и имея физико-математическое описание дополнительных информационных процессов в генетическом аппарате, мы создали аппаратуру, которая моделирует волновые информационные функции живой клетки и ее генетического аппарата. Устройства такого рода являются фактически первыми квантовыми биокомпьютерами. Они позволили осуществить дальнюю многокилометровую передачу генетико-метаболической информации в форме особых электромагнитных полей, введение ее в биосистему-акцептор и стратегическое управление в отношении ее, биосистемы, биохимического и физиологического состояний http://www.self-managing.net/genetica/Zip/SFR.zip. В частности, удалось регенерировать эндокринные железы у животных, а также резко затормозить старение людей.