| Сегодня 31 мая, суббота |
|
|
|
Каталоги Сервисы Блограйдеры Обратная связь Блогосфера
Какой рейтинг вас больше интересует?
|
Почему мы не можем лечить ВИЧ и рак2012-04-30 03:20:00... многих биологов и генетиков, а тем более ... молекулярной биологии и генетики. Для клетки, чтобы ... + развернуть текст сохранённая копия (лингвистико-вероятностно-волновая версия) © Б.И.Бирштейн, А.М.Ярошенко, Гаряев П.П. Леонова Е.А. Тертышный Г.Г. Природа феноменов ВИЧ и рака. Сложности в трактовке. Сущность ВИЧ и рака лежит в той же области, что и сущность Жизни. Но мы до сих пор не понимаем главного в феномене Жизни – как она возникла на Земле и, каким образом она кодируется в хромосоомах. Существует ряд гипотез, каждая из которых, в лучшем случае, лишь часть правды. Отсюда теоретико-биологические трудности в трактовке ВИЧ и рака, а следовательно, и ошибки при их лечении. Оба болезнетворных начала оккупируют главное в любой биологической системе - ее генетический аппарат, то есть сумму “знаний” организма о самом себе. И вот парадокс, вроде бы о хромосомах и ДНК знаем достаточно. Онкогены обнаружены, геном ВИЧ изучен и как эти информационные структуры функционируют в хромосомах понятно. О работе генетического кода и рибосом, тоже вроде бы известно в деталях. Но этого почему-то оказывается недостаточно для разработки абсолютных методов борьбы с раком и ВИЧ. Худшая ложь - это полуправда. В нее привычно верится, особенно, если это полуправда “знания” о генетическом кодировании. Здесь все - неприступный бастион для критики и все догматизировано. Даже основное понятие, стратегическая схема генетического кодирования (ДНКà РНКà Белок) так и названо "Центральная Догма". Атака на эту догму до недавнего времени казалась бессмысленной, обреченной на провал. И как оказалось, напрасно. От того правильно ли мы понимаем генетическое кодирование зависит правильность или неточность стратегии изучения ВИЧ и рака, да и не только этих патологий. Первую брешь в этой догме пробило открытие ревертазы, и догма автоматически превратилась лишь в версию, которая звучит теперь существенно скромнее: ДНКÛ РНКà Белок. Но и эта модификация наших представлений о биосинтезе белков подвергается эрозии, поскольку она - лишь очередное приближение к истине, к пониманию языково-образного плюрализма генома как средству кодирования пространственно-временной структуры биосистем [32, 33]. Что мы хотим доказать? В данном исследовании мы развиваем свои идеи, цель которых не в окончательном разрушении так называемой “канонической” триплетной модели генетического кода, но в развитии и установлении ее точного места в системе знаний о принципах работы хромосом. Да, можно сказать, что триплетный код - истина. Но это истина такого рода, как если бы утверждали, что с помощью алфавита можно написать любое слово. Это верно. Но если мы пытаемся идти дальше, располагая только этим знанием, и доказываем, что с помощью алфавита можно конструировать правильные предложения, то это неверно. Потому, что для построения человеческой речи требуются законы мышления, грамматики и логики. А геном именно речеподобен и логичен, хотя эти фундаментальные черты - не единственный способ выражения его образно-смысловых конструкций. Более того, мы склонны принять идеи В.В.Налимова [43], которые приводят нас к идее, что геном обладает способностью к квазисознанию. Наши логика и модели – это попытка получить знания более высокого уровня о законах построения генетических текстов и иных знаковых структур генома, знания, которые находятся в начальной стадии развития. Основы их были заложены еще в 20-е годы русскими исследователями А.Г.Гурвичем [38], В.Н.Беклемишевым [29] и А.А.Любищевым [41]. Что же можно предложить в развитие и дополнение общепринятой теории генетического кодирования, и как это может помочь решить, как частный случай, проблему ВИЧ и рака? Условно, до полного доказательства, примем три положения, имеющие определенные теоретико-экспериментальные доказательства [8, 32, 33, 37].
Что делать дальше? Допустим, получены окончательные доказательства этих положений. Тогда проблема ВИЧ и рака представляется в совершенно ином интеллектуальном измерении. Попытаемся пояснить эти идеи и факты в теоретико-биологическом и медицинском осмыслении. Например, что такое вещественно-волновой дуализм ДНК и как он связан с многочисленными кодовыми функциями хромосом, существенно отличающимися от известного триплетного генетического кода? В определенном смысле геном выступает как сложный многоволновой лазер с перестраиваемыми частотами. Он излучает свет, гено-знаково промодулированный ДНК по амплитуде, фазе, частоте и поляризации. Более того, геном, вероятно, еще и разер, конвертирующий когерентные знаково поляризованные фотоны в телепортационно связанные с ними когерентные изоморфно знаково поляризованные радиоволны широкого спектра [37] (Гаряев П.П., Тертышный Г.Г., Леонова Е.А. и др. Радиоволновая спектроскопия локализованных фотонов: выход на квантово-нелокальные биоинформационные процессы. Датчики и системы (N9 стр.2-13 за 2000 год).). Геном является также и мобильной меняющейся мультиплексной квазиголограммой, способной при ее многоволновом автосчитывании собственными фотонными излучениями образовывать свето-радиоволновые генознаковые и иные регуляторные структуры [37]. Такие структуры являются реестрами электромагнитных разметочных схем (калибровочных полей) построения пространства-времени биосистем. И наконец, геном – это квазитекстовое образование, с элементами квантовой нелокальности, которое может само себя безинерционно "прочитывать" в миллиардах своих клеток и пользуется полученной при этом информацией как одним из руководств жизнедеятельности и одним из способов организации своей структуры [8, 37]. Наверное, эти идеи о новых информационных измерениях генома для многих биологов и генетиков, а тем более для медиков, воспринимаются сейчас как "китайская грамота". Однако, не для всех из них. Мышление такого рода, пустившее корни в 20 годах в России, набирает силу с резким ускорением в последнее десятилетие. Из этого с определенностью следует, что пока не поздно, необходимо менять стратегию поиска методов лечения ВИЧ и рака, поскольку традиционные попытки решить эти проблемы все более похожи на желание снять богатый урожай, засевая асфальт. Новая стратегия должна быть основана на фундаментальных исследованиях вещественно-волновых и квази-речевых атрибутов генома высших биосистем. Еще раз подчеркнем, что мы рассматриваем хромосомный континуум как знаковый лазерно-радиоволновой излучатель [8, 33, 37]. И этому есть прямые экспериментальные свидетельства. Например, для доказательства лазерных потенций генетических структур мы показали, что in vitro ДНК и хроматин может быть накачан как лазеро-активная среда с последующей лазерной генерацией света [28]. Если мы знаем эти существенные характеристики генома, то возникают естественные и конкретные вопросы – меняется ли знаковый характер лазерно-радиоволновой излучений хромосом при встраивании в них чужеродного генома ВИЧ? И что при этом происходит с "семантикой" излучений при транспозициях онкогенов или любых других мобильных полинуклетидных последовательностей, а такжже при B« Z и иных конформационных переходах в ДНК in vivo? Связано ли это с изменением квази- и голографических программ, то есть, создаются ли новые, изменяются ли старые, “стираются” ли они, и так далее? Динамичен ли при таких перестройках генома параметр поляризации излучений в семиотическом аспекте? Отражается ли все это на работе рибосом? Такие вопросы можно множить и далее. Ответ на любой из них может стать ключевым в трактовке природы ВИЧ и рака. Более подробно о теоретичеких построениях. Или другая проблема, также фундаментальная. Онкогены и геном ВИЧ, занимая определенные позиции в трехмерном пространстве хромосом клеток-хозяев, до какого-то времени не проявляют себя как патогенные факторы. В этом смысле поведение ВИЧ в организме инфицированного человека непредсказуемо. Инкубационный период ВИЧ может варьировать от недели до 10 лет. Считается, что есть определенный механизм индуцирования ВИЧ инфекции из латентного "спящего" состояния, но этот механизм не понят, а следовательно, упускается возможность заставить ВИЧ быть "спящим" в организме человека всегда. Организм и клетки их "не замечают" или даже, как в случае онкогенов, используют в благих целях как факторы роста. Почему они воспринимаемы адекватно (правильно) организмом до времени "X" и семантически перерождаются, вызывая управленческую катастрофу в клетках, после времени "X"? Следуя нашей логике, можно думать, что в патологических состояниях, равно как и в норме, работают, как минимум, четыре фактора - "голографичность" и "лингвистичность" генома, его фоновая (контектная) самоорганизация и его квантовая нелокальность. Эволюция биосистем создала их собственные генетические "тексты" и геном-биокомпьютер как квази-разумный "субъект". Он на своем уровне "читает и понимает" эти тексты. Чрезвычайно важно для обоснования этой элементарной "разумности" генома, что естественные (не существенно на каком языке) человеческие тексты и генетические "тексты" имеют сходные математико-лингвистические и энтропийно-статистические характеристики. Это относится, в частности, к такому понятию как фрактальность распределения плотности частот встречаемости букв (для генетических "текстов" буквы – это нуклеотиды) [21]. Еще одно подтверждение лингвистической трактовки кодовых функций генома получено американскими исследователями [20]. Работая с “кодирующими” и “некодирующими” последовательностями ДНК эукариот (в рамках старых представлений о генах), они пришли к выводу, сходному с нашим и противоречащему догме о том, что знаковые функции сосредоточены только в белок-шифрующих участках ДНК. Они применили метод статистического анализа естественных и музыкальных текстов, известный как закон Ципфа-Мандельброта, а также известный постулат избыточности текстовой информации Шеннона, рассчитываемый как энтропия текстов (относительно энтропии текстов и статистики распределения слов в текстах см., например, [1, 25, 27, 31]. В результате они получили, что “некодирующие” районы ДНК (спейсовые, интронные и другие) более схожи с естественными языками, чем “кодирующие”. Исходя из этого, авторы предполагают, что “некодирующие” последовательности генетических молекул являются основой для одного (или более) биологических языков. Кроме того, авторами был разработан статистический алгоритм поиска кодирующих последовательностей ДНК, который выявил, что белок-кодирующие участки обладают существенно меньшими дальнодействующими корреляциями по сравнению с зонами, разделяющими эти участки. Распределение ДНК-последовательностей оказалось настолько сложным, что использованные методы переставали удовлетворительно работать уже на длинах, превышающих 103 - 102 пар оснований. Распределение Ципфа-Мандельброта для частот встречаемости “слов” с числом нуклеотидов от 3 до 8 показало большее соответствие естественному языку некодирующих последовательностей по сравнению с кодирующими. Напомним, что кодирование авторы понимают как запись информации об аминокислотной последовательности, и только. И в этом парадокс, заставивший их заявить, что некодирующие регионы ДНК - это не просто “junk” (в переводе с английского - “мусор”), а языковые структуры, предназначенные для каких-то целей с неясным пока назначением. Дальнодействующие корреляции в этих структурах авторам также не понятны, хотя и обнаружена нарастающая сложность некодирующих последовательностей в эволюции биосистем. И это продемонстрировано на примере семейства генов тяжелой цепи миозина при переходе от эволюционно низких таксонов к высоким. Приведенные данные [20] полностью соответствует нашим идеям, высказанным ранее и независимо [32, 33], о том, что “некодирующие” последовательности ДНК, а это около 95 - 98 % генома, являются стратегическим информационным содержанием хромосом. Оно имеет материально-волновую природу и поэтому многомерно и выступает, в сущности, как голографическая ассоциативно-образная и одновременно как семантико-семиотическая программа эмбриологического начала, смыслового продолжения и логического конца любой биосистемы. Интуитивно поняв тупиковость старой модели генетического кодирования, авторы [20] с ностальгической грустью прощаются со старой и хорошо послужившей моделью генетического кода, не предлагая, правда, ничего взамен. Омонимо-синонимические неоднозначности генетических текстов. Для чего они нужны организму? Общим фундаментальным семиотико-семантическим свойством естественных и генетических текстов является их синонимии и омонимии. Это обеспечивает для хромосом, также как и для естественных текстов и речи, сверхизбыточность информации, ее многозначность, а поэтому приспособительную гибкость. Многозначность одних и тех же генетических текстов переходит в однозначность за счет эффекта меняющегося положения ДНК-последовательностей в пространстве генома посредством их транспозиций и/или транспозиций их окружения. А это аналог ситуаций в естественных текстах и речи, когда синонимо-омонимические неоднозначности частей семантического поля снимаются контекстом (фоном; фоновый притнцип см. в [44]). В традиционной триплетной модели генетического кода легко обнаруживается омонимии кодирующих дублетов. Значение таких омонимий до сих пор не понято и не оценено, за некоторым исключением [33, 35]. Необъяснимая проблема омонимий кодонов информационных РНК (mРНК) сразу возникла при создании триплетной модели шифровки аминокислот в процессах биосинтеза белков. И сразу стала миной замедленного действия, поскольку правильное объяснение биологического (информационного) смысла таких омонимий автоматически приводит к необходимости существенного уточнения триплетной модели, если не сказать полного ее пересмотра. В чем проявляются омонимии кодонов? Ряд различающихся аминокислот кодируются одинаковыми дублетами в составе кодонов mРНК, а третьи нуклеотиды в кодонах могут меняться хаотически, они "воблируют" (англ. "wobble" – качание) и могут быть любыми из 4-х канонических. Вследствие этого они не коррелируют с кодируемыми аминокислотами [3, 11]. По этой причине возникает семантическая неоднозначность выбора рибосомой антикодонов транспортных РНК (tРНК), несущих аминокислоты. Например, каждый из синонимических кодонов стандартного кода высших биосистем AGT и AGC кодирует серин, а каждый из синонимических кодонов AGA и AGG кодирует аргинин. Поскольку третьи нуклеотиды кодонов mРНК в сочетании со знаковым дублетом не имеют точных аминокислотных коррелятов, а первые два знаковых нуклеотида кодонов одинаковы, но при этом кодируют разные аминокислоты, возникает неоднозначность в выборе антикодонов tРНК. Иными словами, рибосома с одинаковой вероятностью может выбрать и сериновую, и аргининовую tРНК, что может привести к синтезу аномальных белков. В действительности же таких ошибок не происходит, точность белкового биосинтеза чрезвычайно высока. Только в некоторых метаболически аномальных ситуациях происходят ошибки (наличие некоторых антибиотиков, аминокислотный дефицит и др.). В норме рибосома каким-то образом делает правильный выбор антикодонов tРНК из омонимичных дублетов. Мы полагаем, что правильный выбор из дуплетных антикодонов-омонимов реализуется по резонансно-волновым и контекстным (ассоциативным, голографическим) и/или так называемым “фоновым механизмам”. Омонимичность аминокислотного кода может быть преодолена точно так же, как это происходит в естественных языках - путем помещения омонима, как части, в целое, то есть в законченную фразу, контекст которой дешифрует омоним и присваивает ему единственное значение, создавая однозначность. Поэтому mРНК в качестве своего рода “фразы” или “предложения” должна работать в белковом синтезе как функциональное кодирующее целое (нелокально), задающее последовательность аминокислот на уровне ассоциатов аминоацилированных tРНК, которые комплементарно взаимодействуют со всей молекулой mРНК. Макростерические несоответствия между mРНК- и tРНК- континуумами могут сниматься за счет конформационной лабильности макромолекул. При этом роль А-Р участков рибосомы заключается в акцепции таких ассоциатов - предшественников белка с последующей энзиматической сшивкой аминокислот в пептидную цепь. В этом случае будет происходить контекстно-ориентированный однозначный выбор и снятие омонимии дуплет-антикодонов. Можно предсказать в связи с этим, что взаимодействие аминоацилированных-tРНК с mРНК носит коллективный фазовый характер по типу реассоциации (“отжига”) однотяжных ДНК при понижении температуры после “плавления” нативного полинуклеотида. Существуют ли экспериментальные данные, которые можно было бы трактовать в таком духе? Их немало и они сведены в обзорно-аналитическом исследовании [45]. Приведем некоторые из них. Известно, что правильность узнавания молекулами tРНК терминирующих кодонов зависит от их контекстного окружения (подтверждение наших теоретических построений), в частности, от наличия за стоповым кодоном уридина. Так, в работе [9] показано следующее. Вставка строки из девяти редко используемых CUA-лейциновых кодонов после 13-го в составе 313 кодонов тестируемой mРНК сильно ингибируют их трансляцию без явного влияния на трансляцию других mРНК, содержащих CUA-кодоны. Здесь явно проявляются контекстные ориентации трансляции. Хорошо просматривается стратегическая линия влияния строго определенных и далеко расположенных от места образования пептидной связи кодоновых вставок в mРНК на включение или не включение определенной аминокислоты в состав синтезируемого белка. Это дистантное влияние, связанное с континуальностью белкового синтеза (также пример нелокальности функций генетического аппарата), когда mРНК воспринимается белок-синтезирующим аппаратом не только по частям (понуклеотидно, локально), но и как целое (нелокально). Однако, в в цитируемой работе этот ключевой феномен просто констатируется, оставаясь для исследователей непонятным и, видимо поэтому, даже не обсуждается. Таких работ становится все больше. В той, что мы обсуждаем, ссылаются, к примеру, на полдюжины аналогичных результатов, где трактовка в этом смысле также затруднена. Причиной этому, понятно, является несовершенство модели триплетного генетического кода. Модель не точна еще и потому, что существуют необычные протяженные (swollen) антикодоны. Когда они вовлечены в синтез белка, в А-сайте рибосомы участвуют не три, а большее количество пар оснований [45]. Это означает, что догматический постулат триплетности кода нарушается и в этом случае. В [45] приводятся результаты исследований по взаимодействию tРНК-tРНК на рибосоме, и это также полностью подтверждает нашу идею об ассоциате (континууме) нагруженных аминокислотами tРНК как предшественнике белка. В [45] высказана существенная мысль, близкая нам, что эффект действия контекста mРНК на однозначное включение аминокислот в пептид является отражением неких фундаментальных, и пока плохо изученных, закономерностей декодирования генетической информации в процессе белкового синтеза. Напомним, что генетическая информация о синтезе белков занимает всего около 1% объема хромосом. 99% заняты программами существенно более высокого уровня. Прионы – последний удар по центральной догме молекулярной биологии. Как видим, ранние представления о генетическом коде и знаковой работе белок-синтезирующего аппарата носят упрощенный характер. Возможно, последним аргументом в пользу окончательного пересмотра центральной догмы молекулярной биологии служит феномен прионов. Прионы – это низкомолекулярные паразитические белки (PrPsc), поражающие головной мозг животных (бешенство коров) и человека (болезнь Альтсгеймера, синдром Крейцфельда-Якоба и другие). Необъяснимой особенностью прионов является их вирусоподобная штаммоспецифичность. Но штаммоспецифичность присуща только микроорганизмам или вирусам, которые располагают генетическим аппаратом. Вместе с тем считается, что геном у прионов отсутствует, поскольку все попытки обнаружить в составе прионов хотя бы следы ДНК или РНК заканчивались неудачей. Возникает сильное противоречие, которое в очередной раз ставит под сомнение центральную догму молекулярной биологии: генома у прионов нет, а генетические признаки налицо. Не в силах объяснить это, “спасая” центральную догму, все-таки предполагают, что остатки ДНК или РНК прячутся в каких-то складках молекул прионов [10]. Однако, десятилетия исследований прионов, увенчавшиеся нобелевской премией 1997 года Стенли Прузинеру за исследования в этой области, показали абсолютно точное отсутствие в их составе нуклеиновых кислот, а следовательно и генома [23]. Как выйти из этого противоречия? Если придерживаться центральной догмы, то это невозможно. Отказавшись от нее, мы можем представить себе следующий сценарий биогенеза прионов [34]. Главной знаковой фигурой здесь выступает "виртуальный геном прионов", то есть геном временный, взятый заимообразно у клеток-хозяев. Точнее можно сказать, что это белок-синтезирующий аппарат клеток-хозяев. Похоже, прионы в качестве одного из способов размножения сохранили палеогенетический путь, позволяющий им в ряде случаев не пользоваться генами, кодирующими их в хромосомах, а саморазмножаться иным путем, игнорируя центральную догму молекулярной биологии и генетики. Для клетки, чтобы синтезировать прионы, обращаться к их генам - это, хотя и прогрессивный, но организационно и энергетически трудный путь. Прионы могут поступать проще. Мы предполагаем, что NH-группы пептидных связей PrPsc могут реагировать с ОН-группами остатков рибозы акцепторных CCA-последовательностей соответствующих tРНК. В ходе гипотетических ферментативных реакций возникающий поли-tРНК-континуум, колинеарный PrPsc, попарно пространственно сближает антикодоны, образуя ковалентно дискретное “подобие информационной РНК” (пиРНК). Это этап почти обратный синтезу белка на рибосоме. Вероятно, он проходит на А- , Р-сайтах рибосомы. Затем идет синтез РНК на пиРНК. Для этого необходима соответствующая РНК-полимераза, способная работать с ковалентно-дискретной матрицей пиРНК. В этом и заключается "заимообразность" пользования белок-синтезирующим аппаратом клеток-хозяев на время размножения прионов. Такая временность создает иллюзию отсутствия у них генетического аппарата. При этом пептидные цепи прионов служат матрицами, на которых попарно на А-Р участках рибосом, выстраивается поли-tРНК-континуум, образуя дискретные полиантикодоны. Последние, объединяясь попарно, либо сразу служат матрицей для РНК-зависимого синтеза mРНК прионов, либо (в другом варианте) полиантикодоны за счет специфического сплайсинга вырезаются с последующим лигированием в ковалентно непрерывную матрицу mРНК прионов. Далее mРНК прионов на рибосоме полимеризует сами прионы. Это означает, что рибосома работает в обратном направлении и является при этом "прион-полиантикодон-зависимой mРНК полимеразой". И соответтсвенно, в нарушении догмы, информация идет от белка к РНК. Тогда схема догмы окончательно видоизменяется: ДНКÛ РНКÛ Белок. Тогда это уже не догма, а просто рабочая схема, с которой надо работать, уточняя и развивая ее. Штаммоспецифичность прионов, при таком взгляде на их биогенез, объясняется особенностями обратной работы рибосом, временно рекрутируемых при синтезе каждого из штаммов прионов. А эти особенности определяются таксономическим положением биосистем, продуцирующих прионы. Вернемся вновь к пока еще общепринятым основным положениям модели генетического кода: он является триплетным, неперекрывающимся, вырожденным, не имеет “запятых”, т.е. кодоны ничем не отделены друг от друга. Поток информации идет от ДНК к РНК и далее к белку. И наконец, код универсален. Что осталось от этих положений? Фактически ничего. В самом деле, код, видимо, является двух-, трех-, четырех- ... n-буквенным как фрактальное и гетеромультиплетное образование, кодирующее не только отдельные белки, но и функционально связанные ассоциаты белков. Он перекрывающийся за счет сдвигов рамок считывания рибосом. Он имеет запятые, поскольку гетерокодоны могут быть отделены друг от друга последовательностями с иными функциями, в том числе с функциями пунктуации. Код не универсален - в 14 случаях он отличен от стандартного кода высших биосистем. Эти случаи относятся к митохондриальному, дрожжевому, микоплазменному, трематодному и другим кодам низших организмов [5, 6]. И последнее: белок может служить матрицей для РНК, как мы видим на примере прионов. Как понимать реальный генетический, а точнее белковый, код с учетом приведенных противоречий и предлагаемой нами логики рассуждений? Можно постулировать качественную, упрощенную, первичную версию вещественно-волнового контроля за порядком выстраивания аминокислот, задаваемым ассоциатами аминоацилированных tРНК как предшественниками белков. Приняв такую версию, легче понять работу белкового кода как одной из множества иерархических программ вещественно-волновой организации биосистемы. В этом смысле такой код - первый этап хромосомных планов построения биосистемы, поскольку язык генома многомерен, плюралистичен и не исчерпывается задачей синтеза протеинов. Основные положения предлагаемой ориентировочной модели вещественно-волновых знаковых процессов при биосинтезе белков сводятся к следующему:
Таким образом, роль mРНК знаково многовекторна и дуалистична. Эта молекула, как и ДНК, в эволюции знаменует собой узловое событие - взаимодополняющее синергичное единство вещественной и волновой геноинформации. Неоднозначность вещественного кодирования снимается прецезионностью волнового, которое реализуется, вероятно, по механизмам коллективных резонансов и лазерно-голографических (ассоциативных, контекстно-фоновых) эффектов в клеточно-тканевом континууме. Скачок к более развитому волновому регулированию трансляции РНК® Белок сопровождается частичным или полным отказом от правила канонического спаривания аденина с урацилом (тимином) и гуанина с цитозином, свойственного эволюционно ранним и более простым этапам репликации ДНК и транскрипции РНК. Такой отказ информационно необходим, неизбежен и энергетически предпочтителен на уровне высших биосистем. Еще раз подчеркнем, что контекстные ассоциативно-голографические механизмы работы белок-синтезирующей системы организмов теснейшим образом связаны с так называемым “фоновым принципом” [44], а также, вероятно, с многовекторной и многосмысловой логикой управления сложными системами (кенограмматика Герхарда Томаса) [26]. С этой позиции макроконтексты пре-информационных и контексты информационных РНК можно рассматривать как фон, который в данной ситуации и в данной трактовке является "шумовым источником информации". Это обеспечивает резкое усиление сигнала, по которому происходит точный выбор (волновое распознавание) одной из двух омонимичных аминоацилированных tРНК, одна и только одна из которых должна войти в точные белковые “фразу” или “слово”. Этот выбор возможен после выделения когерентной составляющей в форме повторов одних и тех же "осмыслений" (распознаваний) рибосомой одного из двух одинаковых дублетов в кодонах. Ситуацию можно пояснить на простом примере. Скажем, в предложении надо выбрать одно из двух слов (аналогов кодонов с дублетами-омонимами). Эти слова - “суд” и “сук”. Ясно, что выбор зависит от целого предложения, от контекста, который выступает как фон, позволяющий выделить сигнал - нужное слово. Если предложение звучит “я увидел толстый сук на дереве”, то замена здесь слова “сук” на “суд” будет равносильна введению шума и потере сигнала. Вероятно, аналогична роль пре-информационных РНК и интронов - это различные уровни контекстов, которые должны быть каким-то образом “прочитаны” и “осмыслены” живой клеткой и ее рибосомным аппаратом для принятия точного решения по выбору антикодона tРНК в ситуации омонимии. Аппаратом континуального (нелокального) “чтения” контекстных РНК-последовательностей как целого может выступать многоликое семейство солитонов - оптических, акустических, конформационных, вращательно-колебательных и иных, возбуждаемых в полинуклеотиде. Функции таких солитонов могут выступать как способы накопления семантической информации о РНК-контекстах и следующей за этим смысловых регуляций кодон-антикодоновых знаковых взаимоотношений. Смысловые оценки при этом проводятся геномами-биокомпьютерами клеток. В качестве одного из способов континуального прочтения полинуклеотидов можно представить именно солитонный, сканирующий последовательность РНК. Например солитоны бегущих крутильных колебаний нуклеотидов на сахаро-фосфатной оси, физико-математически рассмотренный нами для однотяжных РНК-подобных участков ДНК [30, 36]. Такие солитоны реагируют на изменения последовательностей нуклеотидов модуляциями своей динамики, которая приобретает черты знаковости и может, вероятно, передаваться дистантно, то есть на расстояния, существенно превышающие длину водородных связей. Без дальней (волновой, континуальной) миграции сигнала о целом, то есть о пре-mРНК-mРНК-последовательностях, невозможна реализация ассоциативно-контекстных регуляций синтеза белков. Для этого необходима волновая способность солитонов (а также и голографической памяти) работать не только с частями, но и с целым. Такая континуальность, или что одно и то же, нелокальность, обеспечивает узнавание и правильный выбор рибосомным аппаратом истинного кодона из двух дублет-омонимичных, кодона псевдо-зашумленного фоном (контекстом). Как практиче
Тэги: гаряев, генетика Перспективы волновой генетики2012-04-30 03:05:00... многокилометровую передачу генетико-метаболической информации ... Перспективы волновой генетики // « ... + развернуть текст сохранённая копия Oб авторе Расширенный комментарий к презентацииПолностью презентацию можно посмотреть:
Проведенные исследования, казалось бы, не связаны. Однако это не так. Процесс образования фантомов ДНК был обнаружен нами еще в 1984 г. Обнаружение и регистрация волновых реплик ДНК in vitro подтвердила эти результаты. Это еще раз подтверждает нашу гипотезу о способности ДНК быть структурой волнового автореплицирования и автосканирования внутриклеточного метаболического статуса, а в итоге всего организма с целью биокомпьютерного регулирования жизнедеятельности. Это полностью соответствует основному положению нашей концепции «волнового генома»: хромосомный континуум работает как стратегическая знаковая система на 2-х уровнях — вещества и поля одновременно, реализуя свои функции в качестве квазиразумной системы. Однако это не единственная волновая система саморегуляции многоклеточных организмов. Вторая система основывается на когерентных излучениях хромосом в Уф-, видимой и ИК- областях спектра. Это дает возможность хромосомному аппарату использовать принципы голографической свертки-развертки генетической информации, квантовую нелокальность ее, а также лингвистические особенности работы дуэта «рибосома-иРНК» в рамках понятия квазиразумности генома как биокомпьютера.. Сейчас сложилась парадоксальная и многообещающая ситуация в генетике, молекулярной биологии и, соответственно, в медицине. Изучен геном человека. На это понадобились 10 лет усилий программы «Геном человека», и теперь известна последовательность всех букв в нашем генетическом тексте. Трансгенная инженерия набирает силу, и уже множество растений, животных и бактерий являются носителями искусственно внедренных в них полезных для человека генов. Сделаны первые шаги в клонировании животных с перспективой получить первый клон человека. Но вот парадокс — чем выше наши технологические успехи в генетике и молекулярной биологии, тем дальше мы от всеобъемлющего понимания принципов работы генетического аппарата. Все несомненные успехи в этой области преимущественно связаны с функциями генов, которые кодируют белки. Эти гены занимают около 2% всей генетической памяти хромосом. Остальная, главная часть, т.е. 98%, до сих пор является непонятой генетиками и, вероятно, поэтому они назвали ее «мусором». Есть гипотезы, но все они призваны оправдать существование «мусорной» ДНК как некоего помощника для 2% «кодирующей» ДНК. Либо «мусор» трактуют как «кладбище вирусов»(!) Игнорировать или так наивно понимать роль 98% генома — явная ошибка. Более того, правильно ли мы видим истинную роль 2% генетической информации, если 98% её для нас фактически — «terra incognita»? Понимаем слабо, поскольку не можем полностью излечивать рак, не можем противостоять ВИЧ, не можем бороться против туберкулеза, не можем продлить жизнь людей, по крайней мере, до рубежа 200 лет и т.д. Обещания генетиков обернулись опасными для жизни трансгенными продуктами питания, грозящим дисбалансом биосферы. Клонирование животных дает нам, в основном, тщательно скрываемых уродов, или животных, которые аномально быстро стареют и умирают, как знаменитая овечка Долли. Вполне естественно, что научное сообщество, например, в лице большой группы шведских ученых, начинает бить тревогу по этому поводу http://www.psrast.org/defknthe.htm. Где выход из этого странного состояния обилия экспериментального и противоречивого материала и дефицита достаточно полного теоретического осмысления принципов работы хромосом? Та же группа шведских ученых полагает, что одним из прорывных научных направлений являются наши разработки http://www.rialian.com/rnboyd/dna-wave.doc. Суть наших идей, и что они дают практике, сводится к следующему. Мы исходим из очень простых стратегических соображений. Для успеха в попытках кардинально лечить людей и резко затормозить процессы их старения надо понять языки, на которых клетки общаются друг с другом. В какой-то степени нам удалось это сделать. Оказалось, что это языки тех самых 98% «мусора» в нашем собственном генетическом аппарате http://www.self-managing.net/genetica/Zip/V_Genetica%20(Kniga).zip. 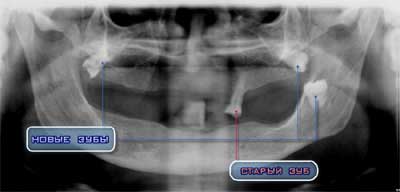 пациентки 60 лет – зубы, выросшие с наклоном под постоянно носимым протезом Основным из них является язык голографических образов http://www.self-managing.net/genetica/Zip/Nestazionarn_golografia.zip, основанный на принципах лазерных излучений генетического аппарата http://www.self-managing.net/genetica/Zip/DNK%20laser.zip, работающего как квазиразумная структура http://www.rialian.com/rnboyd/dna-wave.doc Важно, что нашему генетическому аппарату свойственны реальные процессы, существенно дополняющие триплетную модель генетического кода http://www.self-managing.net/genetica/Zip/Central%20Dogma.zip. В этих процессах используются ранее неизвестные для организмов свойства http://www.self-managing.net/genetica/Zip/FROL.zip и др. Что нам дают эти новые знания? Ответ таков: теперь, зная об этом и имея физико-математическое описание дополнительных информационных процессов в генетическом аппарате, мы создали аппаратуру, которая моделирует волновые информационные функции живой клетки и ее генетического аппарата. Устройства такого рода являются фактически первыми квантовыми биокомпьютерами. Они позволили осуществить дальнюю многокилометровую передачу генетико-метаболической информации в форме особых электромагнитных полей, введение ее в биосистему-акцептор и стратегическое управление в отношении ее, биосистемы, биохимического и физиологического состояний http://www.self-managing.net/genetica/Zip/SFR.zip. В частности, удалось регенерировать эндокринные железы у животных, а также резко затормозить старение людей. Эл.адрес: gariaev@mail.ru Петр Гаряев, д.б.н., акад. РАМТН и РАЕН Перспективы волновой генетики // «Академия Тринитаризма», М., www.trinitas.ru Тэги: гаряев, генетика Фрактальность интеллекта2012-04-30 02:48:00... технологий работы генетиков, медиков. ... волновую искажённую генетико-метаболическую информацию ... + развернуть текст сохранённая копия Гаряев П.П. Oб авторе Насколько я понял, у нас общество взаимного уважения, достаточно узкое, и единомышленников пока мало, но Российская Академия Наук нас уже слушает. Это приятно. Расскажу о том, что мы сделали за последний год. После возвращения из Канады, из славного города Торонто, прошло уже без малого 5 лет, которые были посвящены тому, что мы пытались безуспешно соблазнить медиков, чтобы они среагировали на нас, но, тем не менее, благодаря тому, что я с Аркадием Наумовичем Петровым, наблюдая его работу, поездил по стране, я побывал и в Нижнем Новгороде, где выступал по телевидению. Наши идеи сильно увлекли группу молодых медиков из Нижегородской Государственной Медицинской Академии. Они сказали мне: "Обучите нас, а мы своими руками повторим то, что вы продемонстрировали в Канаде по волновому переносу генетической информации". Говорю им – надо закупить специальный лазер. Они его приобрели, мы его модифицировали, превратив его в квантовый биокомпьютер, который благодаря своей схеме регистрирует знаковую (генетическую) поляризационную динамику биообъектов. Когда фотоны такого биокомпьютера сканируют живые клетки и ткани, то его лазерный луч особым образом поляризуются. Предыстория этого лазера, превращенного нами в биокомпьютер, такова. В 1996 году интуитивно я понял, что один из типов Гелий-Неонового лазера может работать в необычных режимах. Это режимы поляризационного голографирования. Именно они и позволили нам считывать поляризационную, в том числе и генетическую, информацию с живых клеток и тканей. Такую информацию мы стали вводить в самые разные организмы – растения, бактерии и так далее Теоретически мы давно предсказали мощные биологические эффекты при манипуляциях такого рода и много публиковали по этому поводу. Действительно так и получилось. В самом деле, ведь не случайно все мы, живые существа, в основном состоим из оптически активных молекул, в том числе генетических, то есть из ДНК, РНК и белков, способных поляризовать свет. Причем, по разному, в зависимости от свойств и состояний генетических молекул. А эти свойства и состояния и есть генетическая информация, переводимая в поляризационные модуляции фотонов как самим орrанизмом (лазерные излучения хромосом), так и нами in vitro. Организм передает ее поляризующимся лазерным излучением, а потом радиоволнами, во внутри- и внеклеточное пространство, и мы можем повторить этот процесс искусственно. Но реально ли такое повторение? У меня возникла идея, как это сделать. Совестно с моим многолетним коллегой-физиком Георгием Тертышным мы реализовали эту идею. В результате мы научились использовать лазерный пучок для управления метаболизмом организмов. А потом научились использовать превращение биоинформационных фотонов лазера в биомодулированные радиоволны, которые переносили генетическую информацию уже на многие километры и управляли метаболизмом организмов-реципиентов в нужном нам направлении. Так в Канаде, в Торонто мы таким путем вызвали регенерацию поджелудочной железы у крыс in situ на расстоянии около 20 км, но об этом чуть позже. Повторяю, мы разработали не просто новую технологию работы с одним из сотен типов лазеров, но создали квантовый биокомпьютер, имитирующий функции лазерных и радиоволновых излучений хромосом живых организмов. Мы используем лазер в красном диапазоне, а если брать ультрафиолетовый, то биокомпьютер будет на порядки мощнее. Здесь я заканчиваю физико-биологическую часть и перехожу в часть чисто биологическую, точнее генетическую. Есть такое интеллектуальное достижение, которое называется моделью триплетного генетического белкового кода – триумф лауреатов Нобелевской премии Уотсона, Крика, а также Ниренберга и др. западных ученых. Основной вклад в создании модели белкового кода принадлежат Френсису Крику. Он, будучи гениальным человеком, однако, очень скромно оценил свою модель как генетический, ДНК-РНК-овый, способ шифровки последовательностей аминокислот в первичной структуре белков. Но эта модель была быстро и преждевременно превращена во всеобъемлющий генетический и одновременно в торговый брэнд. Может быть, на первых этапах это и было правильно. Полившиеся финансовые потоки в эту область биологии резко ускорили некоторое, более полное, понимание функций генетического аппарата. Почему я заговорил о модели генетического кода, ставшей на десятилетия каноном, не подлежащем критике? Каноны и догмы хороши в религии, но не в науке. Поэтому постепенно модель Ф.Крика из достижения превратилась в тормоз и систему отмывки денег "по научному". В модели кода есть одно ключевое противоречие, которое Крик в 1953г. пытался снять с помощью так называемой "Вобл гипотезы". Она постулирует неоднозначные соответствия кодонов аминокислотам в ген-кодируемых белках и говорит о возможности не канонического и случайного спаривания первого нуклеотида антикодона транспортной РНК (тРНК) с третьим нуклеотидом кодона информационной РНК (иРНК) при ее трансляции в белок [Обозначения: 1-й, 2-й и 3-й нуклеотиды в кодонах и антикодонах – это для упрощения понимания. Тогда 1-й, 2-й и 3-й нуклеотиды в кодонах спариваются с 3-м, 2-м и 1-м нуклеотидами в антикодонах с учетом антипараллельности триплетов. Если быть точным с точки зрения химии полинуклеотидов, то 3-й "воблирующий" нуклеотид в кодоне – это так называемый 3’- нуклеотид, а комплементарный ему 1-й нуклеотид антикодона – это 5’- нуклеотид.]. Проще говоря, при биосинтезе белков иногда реализуется возможность нестрогого соответствия кодон-антикодоновых нуклеотидов в этом положении. Это значит, что образуются неканонические пары оснований [Каноническое термодинамически выгодное спаривание оснований – Аденин/Тимин, Гуанин/Цитозин (для ДНК), Аденин/Урацил, Гуанин/Цитозин (для РНК)], не отличающиеся существенно по геометрическим параметрам (Гуанин-Уридин и др). Кроме того, из Вобл-Гипотезы, да и просто из общей Криковской схемы (модели) кода, автоматически следует, что в кодонах (триплетах) генов только первые два нуклеотида (дублет) кодируют последовательности аминокислот в белковых цепях. Третьи же кодоновые нуклеотиды, не участвуют в кодировке аминокислотных последовательностей в белках. Эти третьи, хотя и детерминированы жестко молекулой ДНК, но допускают произвольные, случайные, не канонические спаривания с первыми нуклеотидами антикодонов транспортных РНК, переносящих аминокислоты. А посему эти первые нуклеотиды антикодонов могут быть любыми из 4-х возможных. Следовательно, третьи нуклеотиды в кодонах и спаривающиеся с ними первые в антикодонах, не имеют знакового характера и играют роль "стерических костылей", заполняющих "пустое место" в кодон-антикодоновых связках. Короче говоря, первый нуклеотид в антикодонах случаен, "воблирует" - от английского ‘wobble’ (качание, колебание, виляние). Это очень важно и скажу почему. Поскольку всего 64 кодона, а аминокислот 20, то получается избыток кодонов, и соответственно кодирующих дублетов. Этот избыток, используя лингвистическую аналогию, называют синонимией, то есть несколько дублетов кодируют одну и ту же аминокислоту, отсюда понятно существование изоакцепторных транспортных РНК (тРНК), переносящих аминокислоты на А-сайт рибосом. Такая синонимическая щедрость даже хороша – это избыточность информации. Но при белковом синтезе неизбежно возникают ситуации, когда одинаковые дублеты в кодонах информационной РНК, вынуждены как-то "кодировать" разные аминокислоты и/или стоп кодоны. Напомню, что третьи и первые нуклеотиды в кодон-антикодоновой "костыльной" связке находятся вне знаковой "игры", а это порождает лингвистическую неоднозначность, то есть ситуацию омонимии. Как, например, для слова "коса", когда смысл его неоднозначен. И соответственно, перед рибосомой, читающей одинаковые (омонимичные) дублеты кодонов, встает задача выбора из двух разных аминокислот, переносимых тРНК, одной единственной "правильной" аминокислоты и/или стоп-сигнала. Если выбор неправилен – белок получится аномальным, что может привести к метаболической катастрофе и гибели организма. Как же происходит правильный выбор? А он, как установлено, правильный и на 99,999% точен. Рибосома практически не ошибается, а казалось, должна бы, если автоматически следовать модели кода! Модель Ф.Крика это не объясняет. И сам Ф.Крик это видел и признавал в своих, опубликованных незадолго до смерти, воспоминаниях, но ограничился фиговым листочком Вобл Гипотезы. И вся последующая генетика никак не комментировала эту зияющую дыру в модели кода: модель противоречива, не точна, но синтез белков, тем не менее, безошибочен. И только в 1978г. швед Ульф Лагерквист первым открыто заявил об этом противоречии в модели кода [Lagerkvist U., 1978, “Two out of Three”: an alternative method for codon reading. Proc. Natl. Acad. Sci. USA, v.75, p. 1759-1762.]. Но и Лагерквист также не предложил ничего внятного в объяснение, не обострил проблему, а лишь констатировал ее. Всех устраивало, что синтез белков – точный процесс и требовать от Криковской модели большего, вроде как, и не требовалось. "Жена цезаря вне подозрений". А потом появились работы о так называемых контекстных ориентациях при трансляции иРНК рибосомами. Это был момент истины. Было экспериментально показано, что выбор аминокислот рибосомой в омонимичных и других ситуациях зависит не только от дублетов кодонов, но и от контекста иРНК. Точно как в человеческой речи (текстах), когда, например, упомянутый омоним "коса" обретает точный смысл только в контексте целой фразы или предложения. Сей фундаментальный факт биологи опять-таки только констатировали. И все. Никакого анализа, почему так происходит и что это значит. Меня такое положение дел не устраивало. Думаю, и многих других биологов. Это заговор молчания научного официоза. А причина проста. Анализ поведения генетического аппарата в ситуации омонимий логически приводит к сильному, я бы сказал, фундаментальному тезису о квази разумности генома. Для правильного выбора аминокислоты в ситуации омонимической неоднозначности белок синтезирующей системе необходимо ПРОЧИТАТЬ, прочитать не в метафорическом смысле, но реально, всю молекулу иРНК, то есть контекст и ПОНЯТЬ его смысл. И ПРИНЯТЬ РЕШЕНИЕ о выборе "правильной" аминокислоты или стоп-сигнала. Это можно сделать только С ПОМОЩЬЮ АКТА МЫШЛЕНИЯ, РАЗУМНО, что означает наличие у генома МЕНТАЛЬНОГО НАЧАЛА. Следовательно, геном базируется не только на чистой биохимии и физике взаимодействий информационных биомакромолекул, что стыдливо умалчивалось официальным научным сообществом в угоду жёсткому материализму и меркантильным соображениям. Понятие квази разумности генома выводит в совершенно другие области знания, которые сейчас пока либо недоступны, либо намеренно прикрываются. Одновременно это и пересмотр мировоззренческих позиций в отношении функции и происхождения наших хромосом, а заодно, пересмотр всех технологий работы генетиков, медиков. Следовательно, это ведет к переориентировке финансовых потоков в биологии и медицине, что многим не по вкусу. В дополнение скажу, что недавно в работах В.И.Щербака доказан ещё один фундаментальный факт – белковый код оперирует математикой с использованием запредельно абстрактного понятия НУЛЯ [Shcherbak V.I., 2003, Arithmetic inside the universal genetic code. BioSystems, v.70, pp.187–209.]. В.И.Щербак обнаружил в системе генетического исчисления использование функций ноля. Это чрезвычайно важное обстоятельство, поскольку ноль – сугубо мыслительное порождение, дающее начало координатному сознанию с его количественными мерами оценки внешнего мира, которые интерпретируется внутренним организменным генетическим сознанием-исчислением. Таким образом, цифры (наряду с буквами) становятся неотъемлемой частью генетического (белкового) кода. И мы должны принять очередное сильное положение, что in vivo существует арифметическое управление в лингвистической и/или текстовой генетике. Итак, белок синтезирующая система и геном в целом обладают способностью к квази мышлению и математическому исчислению как части его. Замечу, что это не мышление уровня головного мозга, а квази мышление генетического аппарата как биокомпьютера. Здесь мы имеем дело с разными фрактальными размерностями одного и того же феномена – мышление и сознание на уровне коры головного мозга (одна размерность высшего порядка) и мышление и сознание на уровне функций хромосом и белок синтезирующей системы (другая размерность того же, но микро масштаба). Эти простые соображения выводят биологию, генетику, медицину в совершенно другие ареалы, и нам ясно, что каноническая модель генетического кода явно НЕ ПОЛНА. Геном – это лингвистико-математическая структура, тексты и алгоритмы Творца. При этом надо учесть и наши последние теоретико-экспериментальные разработки. В них показано, что генетические тексты и иные образные знаковые структуры генома могут храниться в форме поляризационных голограмм в жидких кристаллах хромосом [Тертышный, Гаряев, 2007, Волновые генетические нанотехнологии управления биосистемами. Теория и эксперименты. Новые медицинские технологии, 2007, №7, с.49-64]. Этот информационный пул огромен и занимает большую часть генома – около 98%. Удивительно, эти 98% генетического материала не поняты официальной генетикой и названы "мусором"! Но это тема для отдельного большого разговора. Существенно также и то, что высшие формы сознания человека на уровне коры головного мозга реализуются, видимо, также хромосомным аппаратом, как основной информационной фигурой любых клеток. Коррелятом мышления и сознания опять-таки выступают текстовые структуры быстро синтезирующихся фракций белков. Замечу, что самый мощный синтез белков имеет головной мозг, нейроны коры мозга. Работа генома мозга идет также по пути голографирования с использование ресурсов ДНК. Таким образом, идеальное начало мышления-сознания базируется на материальных структурах информационных молекул ДНК, РНК и Белков, которые являются отображениями речи и технологий Творца, как инобытие Его Мысли. Вернёмся к официальной генетике. Мы видим с её стороны полное игнорирование очевидных зияющих дыр в понимании работы генома, следовательно, и организма человека, животных, растений, бактерий, вирусов. К чему это ведёт и уже привело? Взять, к примеру, многомиллиарднодолларовую международную программу Геном человека. За теоретическую основу её взята Криковская модель генетического кода. Итог 15-летней программы таков – изучен порядок следования 3 миллиардов нуклеотидов в ДНК хромосомного набора человека. Найдено около 30000 генов. Это около 2% от всей ДНК генома человека. Остальная, главная часть, то есть эти 98%, научный официоз назвал мусором! Установлено, по этим генам человек практически неотличим от собственной кишечной палочки, от ослов, свиней, тем более обезьян. В основном это гены ферментов, которые, как рабочий инструментарий метаболизма, универсален почти для всех живых существ. Извините, господа генетики, я не согласен. Генетически мы существенно иные по сравнению с бактериями и свиньями! И отличия эти в иных принципах генетического кодирования, которые мы, и не только мы, обнаружили, и которые реализуются в тех самых 98% "мусора" геномной ДНК. А вы, господа официальные генетики и молекулярные биологи, об этом знать не хотите и объявляете лженаукой. Ну не вредительство ли? Программа Геном Человека впустую потратила огромные деньги, фактически отмыла их, обеспечив на многие годы безбедной жизнью кланы паразитирующих на незнании реальных псевдоученых. Но это полбеды. Хуже другое – неправильное, неполное понимание принципов генетического кодирования привело нас к проклятию генной "инженерии", когда тиражируются вирусные, бактериальные, растительные, животные химерные монстры, грозящие генетическим коллапсом всей планеты. Что же предлагаем мы, пытаясь найти научные тропы к таким ясновидцам России, как Грабовой, Петров, Арепьев и их ученики? И что нас объединяет с ними? ДНК – это речь Творца и построены тексты ДНК сходно с человеческой речью. Много работ на эту тему опубликовано, особенно израильтяне много сделали. Это относится не только к известным генам, но мы полагаем, что голограммы генома в тех самых пресловутых 98% "мусора" содержат огромное количество интерференционно запакованных генов. Вот вам пример, косвенно подтверждающий эту мысль. Недавно опубликована статья группы Прюита из Соединенных Штатов [Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).], причем не где-нибудь, а в Nature – нобелевский журнал! Так эта работа заставила вздрогнуть всю официальную менделевскую генетику. Как показали изящные эксперименты, скромное растеньице, ‘Резушка Таля’ (Arabidopsis thaliana), способно "вспомнить" и выставить напоказ предковый ген HotHead, который отсутствовал в его геноме. При этом HotHead (нормальный ген) заменял мутантный hothead. Полезная и многообещающая эндогенная манипуляция… Геном исходных мутантных семян предварительно анализировали блоттингом и секвенированием. Не было гена, а во взрослых растениях из этих семян ген HotHead выскакивал, как чёрт из коробочки! Откуда он взялся? Генетика до сих пор чихает от этого результата, а Мендель, наверное, перевернулся в гробу. Шум в Интернете среди научного официоза стоит страшный. Кстати, мы аналогичный результат ухода от мутаций получили за 2 года до американцев и опубликовали, но, увы, не в Nature, а посему он не замечен [П.П.Гаряев, Е.А.Леонова, 2003, Странный мир волновой генетики. Журнал "Сознание и физическая реальность", т.8, №6, с.27-40.]. Объяснение этому феномену простое – предковые гены исчезают, но не бесследно, они остаются в памяти генома, упакованные в голограммы "мусорной" ДНК. Если такие гены полезны организму, он их извлекает. Такие извлечения бывают аномальными и называются атавизмами. Однако я отвлекся. Так что же нас объединяет с ясновидцами, лечащими болящих СЛОВОМ и собственными излучениями? Связь очевидна – СЛОВО ЕСТЬ КВАЗИГЕНЕТИЧЕСКИЙ МАТЕРИАЛ на уровне микро- и макросоциальных отношений. Словом и лечат, и убивают. Разновидность такого рода воздействий на людей достаточно хорошо известна. Это нейро-лингвистическое программирование (НЛП). Кто только им не пользуется – от нашей мамы в детстве до таких говорунов как Жириновский. Но если лечащий, кроме искусства слова, владеет еще и повышенной по сравнению с нормой способностью излучать знаковые физические поля, то есть те, что излучают хромосомы, клетки и голова человека, то эффекты могут быть весьма значительными. Что мы и видим, начиная от воскрешения Христом Лазаря, кончая регенерацией органов и тканей у больных, вызываемые кудесниками ясновидения. Излучательная часть феноменов такого рода особенно близка нам. Мы исследуем генетический аппарат не только как текстовую структуру, но как структуру, излучающую реальные, управляющие организмом физические поля – звук, свет, радиоволны. Этими полями мы научились оперировать как лечебными. Одним из спектров электромагнитных полей, излучаемых хромосомами, точнее, их ДНК, является лазерный свет от ультрафиолета до ИК диапазона. Фактически, это предсказал наш ученый А.Г.Гурвич в 20-х годах прошлого века. Он же автор теории биологического поля. Именно его исследования стимулировали работу моей группы, начиная с 1985 года. Теорию А.Г.Гурвича предельно кратко можно выразить одной фразой – хромосомы имеют волновой эквивалент. Мне посчастливилось в течение 10 лет, с 1987 по 1997 годы, поработать в Физическом Институте АН среди талантливых физиков-лазерщиков и теоретиков. Помню, под влиянием идей Гурвича, я предложил группе учёных ФИАНа сделать лазер на молекулах ДНК и на хромосомах. Встречено было скептически, но убедил, и такую работу мы сделали – получили эффект лазерной накачки ДНК и хромосом in vitro с помощью пикосекундного лазера на парах меди. Это было опубликовано в журнале "Квантовая Электроника" в 1996 г. [А.М. Агальцов, П.П. Гаряев, В.С. Горелик, И.А. Рахматуллаев, В.А. Щеглов, 1996, Двухфотонно-возбуждаемая люминесценция в генетических структурах. Квантовая электроника, v.23, N2, с.181-184.] А спустя 6 лет близкие результаты получили японские авторы [Y. Kawabe, L. Wang, T. Nakamura, and N. Ogata Thin-film lasers based on dye-deoxyribonucleic acid-lipid complexes Applied Physics Letters -- August 19, 2002 -- Volume 81, Issue 8, pp. 1372-1374.]. Теперь уже никто не мог возразить, что хромосомы могут работать как лазерный источник и при этом фотоны его могут передавать генетический сигнал от клетки клетке. Насчёт фотонной генетической передачи, это было гипотетично, хотя имелись кое-какие работы в этом направлении – исследования Дзян-Каньдженя, Казначеева, Бурлакова, Будаговского. Это наши, Россияне. Американцы тоже думали в этом направлении – Ричард Миллер в 1973 году первым предложил идею голографической памяти хромосом. Все это было, но не четко, без развитой теории и с малым запасом экспериментов. И вот в 1994 году мы совместно с Георгием Тертышным в Институте Проблем Управления РАН начали длительный цикл исследований, целью которых было создание аппаратуры, моделирующей знаковые (генетические) излучения хромосом в организме. В 1996г. мне пришла в голову идея, что основной пул генетической информации записан в основном на уровне оптической активности ДНК хромосом в голографическом варианте, на способности вращать плоскость поляризации падающего на них света. Хромосомный континуум многоклеточного организма является поляризационной голограммой и одновременно лазером, считывающим генетическую световую информацию с самого себя. Чтобы доказать это, я предложил использовать и модифицировать специальный лазер, чувствительный к оптической активности хромосом и, соответственно, к информации, записанной в хромосомах на этом уровне. Начали экспериментировать. Одна из серий опытов была особенно убедительна. Мы взяли чистую ДНК, выделенную из растения Arabidopssis thaliana. Препарат ДНК – это всегда испорченный генетический материал, это не целостные неповрежденные хромосомы. ДНК повреждается при выделении, удаляются специальные хромосомные белки. Вот такую испорченную ДНК-матрицу мы сканировали пучком нашего лазера. Такой тип лазера, как оказалось, осуществляет переход фотонов в радиоволны. О переходе этом тоже особый длинный разговор. Мы дали физико-математический формализм этому явлению. Эти радиоволны информационно связаны с фотонами лазерного луча. И если луч зондирует препарат ДНК, то информация, считанная с препарата ДНК, переписывается на радиоволны. А те, в свою очередь, переносят её на неопределенно большие расстояния во все четыре стороны. Вот мы и попробовали в 2000 году передать такую волновую генетическую и, напомню, поврежденную генетическую информацию с ДНК арабидопсиса на расстояние около 6 км между нашим Институтом Проблем Управления РАН и Институтом Общей Генетики РАН. Что из этого получилось? А получилось то, что мы и ожидали. Проростки семян арабидопсиса-реципиента приняли передаваемую ложную информацию и мутировали. Сотни проростков мутировали. Чисто фенотипически это были так называемые летальные эмбриональные мутации по Мюллеру. Но мутации необычные, не связанными с разрывами ДНК в хромосомах реципиентов (энергия радиоволн мала). Во 2-м поколении мутации наследовались, а в 3-м ушли. Что это были за необычные мутации, вернее, квазимутации, нам до сих пор неясно. Вероятно, они были связаны с наведенными изменениями топологии хромосом растений, хромосом как динамичных жидких кристаллов (голограмм). Это был уход от мутаций и возврат к исходному фенотипу растений, аналогично тому, что наблюдала группа Прюита с геном HotHead, о чем уже говорил. Потом аналогичный квазимутагенез и уход от него в 3-м поколении мы увидели на растении картофеля. Подчеркну адресность воздействия – только близко родственные реципиенты воспринимали волновой морфогенетический сигнал. Что означали такие, казалось бы, нехорошие эксперименты? Ведь мы повреждали геном реципиентов. Он подсказал нам направление будущих лечебных и прочих полезных воздействий на генетический аппарат такими радиоволнами. Если мы вызвали подобие неких мутаций у пространственно далеко расположенного реципиента, то нельзя ли таким образом избирательно уничтожать сорняки, насекомых-вредителей и переносчиков болезней, патогенные бактерии и вирусы и так далее, и тому подобное. Открылись широкие горизонты. Любая хорошая идея должна быть простой в своей основе. Наша идея очень простая, но обросла гигантским физико-математическим аппаратом. Когда вы возьмёте наши статьи, поймёте, что не так всё просто, хотя основа ясна. Эта основа называется "делай как я". Если вы даёте волновую искажённую генетико-метаболическую информацию на объект воздействия, на реципиент, то получаете фактически адресное управление метаболизмом. Сейчас о другом. Регенерация органов и тканей – то, что является главным в работе Аркадия Наумовича Петрова и то, что делают другие кудесники России. А нельзя ли нам регенерировать органы и ткани у людей нашей технологией? Это было бы просто здорово и подвинуло бы медицину в этом направлении, а также поддержало бы лечащих ясновидцев с позиции науки. И было бы великолепным подтверждением, что ментальные технологии ясновидцев базируются не на бреде сивой кобылы в лунную ночь, а основаны на классической физике и волновой генетике. Решили попробовать. Есть так называемая аллоксановая модель диабета. Она достаточно изучена. Хорошо известно, что в методы лечения диабета вложены миллиарды долларов. Толку чуть. Инсулин – основное средство. А мы решили попытаться вырастить поджелудочную железу прямо в организме, использовав аллоксановую модель. Есть такой мощный яд – аллоксан. Достаточно крысе инъецировать 50 мг аллоксана, она гибнет от диабета – разрушаются бета клетки и инсулин исчезает из крови, что ведёт к гибели животных. Первый эксперимент поставили в ИПУ РАН в 2000 году совместно с Институтом Теоретической и Эксперимнтальной Биологии в Пущино. Эксперимент был удачным, крысы выздоровели, поджелудочная железа регенерировала. Но крыс было мало – 36, поэтому со статистикой не проходило. Однако мы были замечены в Канаде и получили приглашение поработать именно по этой теме. Прилетели в Торонто и с 2001 по 2002 год работали в мощной фирме SynX. Там были идеальные условия для работы. Мы повторили наши положительные результаты на существенно большем числе крыс с хорошей статистикой. При этом продемонстрировали, что расстояние не играет критической роли для передачи волнового морфогенетического сигнала, способного запустить процессы регенерации поджелудочной железы in situ – прямо в теле животного. Расстояния от лазера до больных животных составляло от 1 см до 20 км. Результаты вызвали шок у канадских исследователей, которые участвовали в экспериментах. Эксперименты для нас были мгновенно свернуты. Нам сказали, что выздоровевших крыс усыпили! На вопрос – почему? Ответ был – эксперименты закончены. После этого мы вернулись в Москву, сами понимаете, в каком состоянии. Наши попытки возобновить эти работы в столице не встретили никакой поддержки, более того, академик Кругляков публично объявил меня лжеучёным, приводя в пример "бредовость" наших идей относительно создания лазера на молекулах ДНК. Я ему ответил открытым письмом в Интернете. А публикация японцев, подтвердивших наши результаты по ДНК-лазеру, поставила жирную точку на Круглякове. Словом, с 2002 по 2007 годы я пытался продолжить исследования. Безуспешно. А в ИПУ РАН меня уже не пустили, спасибо Круглякову. Но судьба вывела меня на Аркадия Наумовича Петрова. Начались поездки по городам России, выступления, где я обосновывал научную состоятельность направления работ групп Грабового и Петрова, а заодно рассказывал о наших исследованиях. И нашлась-таки группа медиков, среагировали, попросили их обучить. Я им всё рассказал, обучил. Эти ребята полностью повторили канадские эксперименты, это опубликовано в ВАК-овском журнале [Гаряев П.П., Кокая А.А., Мухина И.В., Леонова-Гаряева Е.А., Кокая Н.Г., Влияние модулированного биоструктурами электромагнитного излучения на течение аллоксанового сахарного диабета у крыс. Бюллетень Экспериментальной Биологии и Медицины, №2, с.155-158 (2007)]. Удивительно, официальная наука позволила публикацию по волновым генам. Значит, какая-то подвижка имеется. Вслед за этой статьей опубликовали большой теоретический анализ полученных результатов [Тертышный Г.Г. Гаряев П.П., 2007, Волновые генетические нанотехнологии управления биосистемами. Теория и эксперименты. Новые медицинские технологии, 2007, №7, с.49-64], и одна в продолжение этого анализа еще предстоит. ВОПРОСЫ – Когда выйдет Ваша следующая третья книга? П. Гаряев: Она практически готова, последние штрихи остались. – У нас в Алма-Ате съедается червями яблоневый сад, то есть эту проблему тоже можно решить? П. Гаряев: Так же, как мы вызывали мутации у арабидопсиса и на картофеле. Нужен препарат клеточных ядер или ДНК этих червей. Далее мы модифицируем этот материал рентгеном или ультрафиолетом. Это и будет источник ложной волновой информации для червей, которая дестабилизирует нормальную работу их генома. Черви погибнут. – Как Вы подошли к теории Аркадия Наумовича? Как вы стыкуетесь? А. Петров: Да я поругался в Интернете в их адрес, а он услышал. П. Гаряев: И правильно сделал, что ругался. Я увидел в Интернете его критический комментарий в отношении безопасности наших работ. Пришел к Аркадию Наумовичу, объяснил, постарался убедить, что работы по волновой генетике будут развиваться независимо от того, хотим мы этого или нет, а также высказал соображения, что именно эти работы могут дать реальное научное обоснование ясновидению, понимание, как геном человека отвечает на речевые и ментально-волновые воздействия, исходящие от ясновидящего. С этого всё и пошло в наших контактах. А. Петров: А когда эти процессы видишь в режиме ясновидения, видишь процессы сжатия, компрессии ДНК информации, то становится ясно, что в ДНК есть те же ситуации, о которых Пётр Петрович говорил. И когда хромосомная ДНК высвобождает свою волновую информацию, то она работает как голограмма, давая речевые и иные образы. При этом часть ДНК может читаться напрямую, когда она, как жидкий кристалл, составлена плотной упаковкой её нитей-текстов. И такие тексты могут читаться не только вдоль ее цепочек, но и поперек, справа налево, слева направо, вверх, вниз, вообще многомерно, по всем направлениям. Это огромная информация. И она, конечно же, имеет цифровой эквивалент. П. Гаряев: Это как кроссворд, только он многомерный. А. Петров: Есть ещё очень интересная фаза – когда поэтические тексты начинают оттуда выходить. Когда достигаешь определённого погружения в собственное сознание, анализируя ДНК-континуум, оттуда выходит очень значимая, но поэтизированная информация, то есть она рифмованная. Оказывается, что и такая возможность есть. При этом мы знаем, что разные народы пишут по-разному (справа налево, слева направо и так далее), то есть получается, что письменность и структуры ДНК имеют некую общность. То есть каждый народ отрабатывает какую-то глобальную программу, которая имеет фиксацию на уровне ДНК. Пётр Петрович со стороны науки, а мы с другой стороны, видим одно и то же. И мы также давно работаем со стволовыми клетками, регенерация именно на основе стволовых клеток ведётся. П. Гаряев: Какая сейчас перспектива возникает. Вот этот широкополосный электромагнитный спектр ДНК, что мы извлекаем из нее. С ним можно оперировать по определенным программам и достигать самых разных биологических эффектов. Только осторожно надо работать. Тут непочатый край работы. Это тоже будет своего рода волновая генная инженерия. Только мы не будем идти напролом, как идут сегодняшние трансгенные "инженеры". Разобраться сначала надо в работе генома, а уж после манипулировать им на благо людей и всего живого на Земле. Спектры, что мы регистрируем, можно получать не только с ДНК, клеток и организмов. Любое вещество дает эти необычные и неизвестные ранее спектры. Вообще это новый тип динамической поляризационно-лазерно-радиоволновой спектроскопии. Мы тут в самом начале. Например, можно получить спектры камней. Многие из них обладают биологической активностью, а спектры усиливают её во много крат. И все они дают свои уникальные мелодии, если перевести спектр из радиоволн в звук. А. Петров: В своё время, нам привели мальчика, когда мы с Игорем Витальевичем Арепьевым работали, и у него была кризисная ситуация. И одновременно было видно, есть прогнозная фаза по будущему, что если сейчас выводишь ребёнка из этого кризиса, он дальше становится серийным убийцей, маньяком. И вот здесь разница между прибором и специалистом, который всё-таки развивает себя, а не прибор. Потому что он тогда имеет возможность прогнозировать будущее и знать, что произойдёт дальше. И по мальчику было найдено решение – сначала его определённое духовное развитие, и после этого – коррекция его здоровья. И тогда вот эта тяжелая прогнозная фаза ушла. П. Гаряев: Подводя итог, вернемся к нашим экспериментам с крысами. Фактически, мы программировали стволовые клетки. Эта задача сейчас стоит перед медициной, поскольку программирование стволовых клеток – это возможность заменять старые и больные органы у людей на новые, затормозить их старение, продлить активную жизнь. Сюда вкладывают во всем мире миллиарды долларов. И, надо сказать, впустую. Без понимания принципов волновой генетики эта проблема станет черной дырой, куда впустую будут вбрасывать деньги и труд исследователей. Вопрос только – зачем? Ведь решение проблемы – вот оно, преподносится на блюдечке. Но это блюдечко тяжело далось – это результат нашей 20-летней теоретической и экспериментальной работы. Приношу благодарность моим многочисленным соавторам и коллегам, с которыми я эти десятилетия работал, и без сотрудничества с которыми ничего бы не получилось. Особенно я благодарен физикам Оптического Отдела ФИАНа, с которыми мне удалось плодотворно поработать на протяжении 10 лет, а также моему наиболее близкому коллеге, блестящему физику – Георгию Георгиевичу Тертышному. www.trinitas.ru Тэги: гаряев, генетика, филология Ученые обнаружили новый способ регенерации человеческих конечностей2012-04-28 23:59:15Гекконы могут частично восстанавливать свой хвост, морские звезды свои многочисленные конечности, но ... + развернуть текст сохранённая копия Гекконы могут частично восстанавливать свой хвост, морские звезды свои многочисленные конечности, но человек не обладает этой удобной возможностью. Саламандры вообще являются единственными позвоночными, которые могут регенерировать целые конечности. Тем не менее, человек может частично восстанавливать повреждения печени или даже кончики пальцев. Новое исследование показывает, что эта способность может быть улучшена. Исследование, которое финансируется за счет Министерства обороны [...] Тэги: биоинженерия, генетика, медицина Ученые обнаружили новый способ регенерации человеческих конечностей2012-04-28 23:59:15Гекконы могут частично восстанавливать свой хвост, морские звезды свои многочисленные конечности, но ... + развернуть текст сохранённая копия Гекконы могут частично восстанавливать свой хвост, морские звезды свои многочисленные конечности, но человек не обладает этой удобной возможностью. Саламандры вообще являются единственными позвоночными, которые могут регенерировать целые конечности. Тем не менее, человек может частично восстанавливать повреждения печени или даже кончики пальцев. Новое исследование показывает, что эта способность может быть улучшена. Исследование, которое финансируется за счет Министерства обороны [...] Тэги: биоинженерия, генетика, медицина
Главная / Главные темы / Тэг «генетика»
|
Категория «Новости»
Взлеты Топ 5
Падения Топ 5
Популярные за сутки
300ye 500ye all believable blog cake cardboard charm coat cosmetic currency disclaimer energy finance furniture hollywood house imperial important love lucky made money mood myfxbook new poetry potatoes publish rules salad sculpture seo size trance video vumbilding wardrobe weal zulutrade агрегаторы блог блоги богатство браузерные валюта видео вумбилдинг выводом гаджеты главная денег деньги звёзды игр. игры императорский картинка картон картошка клиентские косметика летящий любить любовь магия мебель мир настроение невероятный новость обзор онлайн партнерские партнерских пирожный программ программы публикация размер реальных рубрика рука сайт салат своми стих страница талисман тонкий удача фен феншуй финансы форекс цитата шкаф шуба шуй энергия юмор 2009 |
Загрузка...
| Copyright © 2007–2025 BlogRider.Ru | Главная | Новости | О проекте | Личный кабинет | Помощь | Контакты |