| Сегодня 7 апреля, вторник |
|
|
|
Какой рейтинг вас больше интересует?
|
Главная /
Каталог блоговCтраница блогера My personal blog./Записи в блоге |
|
My personal blog.
Голосов: 1 Адрес блога: http://abdullin.blogspot.com/ Добавлен: 2010-01-26 23:41:06 блограйдером rusfbm |
|
Фрактальность интеллекта
2012-04-30 02:48:00 (читать в оригинале)Гаряев П.П.
Oб авторе
Насколько я понял, у нас общество взаимного уважения, достаточно узкое, и единомышленников пока мало, но Российская Академия Наук нас уже слушает. Это приятно.
Расскажу о том, что мы сделали за последний год. После возвращения из Канады, из славного города Торонто, прошло уже без малого 5 лет, которые были посвящены тому, что мы пытались безуспешно соблазнить медиков, чтобы они среагировали на нас, но, тем не менее, благодаря тому, что я с Аркадием Наумовичем Петровым, наблюдая его работу, поездил по стране, я побывал и в Нижнем Новгороде, где выступал по телевидению. Наши идеи сильно увлекли группу молодых медиков из Нижегородской Государственной Медицинской Академии. Они сказали мне: "Обучите нас, а мы своими руками повторим то, что вы продемонстрировали в Канаде по волновому переносу генетической информации". Говорю им – надо закупить специальный лазер. Они его приобрели, мы его модифицировали, превратив его в квантовый биокомпьютер, который благодаря своей схеме регистрирует знаковую (генетическую) поляризационную динамику биообъектов. Когда фотоны такого биокомпьютера сканируют живые клетки и ткани, то его лазерный луч особым образом поляризуются. Предыстория этого лазера, превращенного нами в биокомпьютер, такова. В 1996 году интуитивно я понял, что один из типов Гелий-Неонового лазера может работать в необычных режимах. Это режимы поляризационного голографирования. Именно они и позволили нам считывать поляризационную, в том числе и генетическую, информацию с живых клеток и тканей. Такую информацию мы стали вводить в самые разные организмы – растения, бактерии и так далее Теоретически мы давно предсказали мощные биологические эффекты при манипуляциях такого рода и много публиковали по этому поводу. Действительно так и получилось. В самом деле, ведь не случайно все мы, живые существа, в основном состоим из оптически активных молекул, в том числе генетических, то есть из ДНК, РНК и белков, способных поляризовать свет. Причем, по разному, в зависимости от свойств и состояний генетических молекул. А эти свойства и состояния и есть генетическая информация, переводимая в поляризационные модуляции фотонов как самим орrанизмом (лазерные излучения хромосом), так и нами in vitro. Организм передает ее поляризующимся лазерным излучением, а потом радиоволнами, во внутри- и внеклеточное пространство, и мы можем повторить этот процесс искусственно. Но реально ли такое повторение? У меня возникла идея, как это сделать. Совестно с моим многолетним коллегой-физиком Георгием Тертышным мы реализовали эту идею. В результате мы научились использовать лазерный пучок для управления метаболизмом организмов. А потом научились использовать превращение биоинформационных фотонов лазера в биомодулированные радиоволны, которые переносили генетическую информацию уже на многие километры и управляли метаболизмом организмов-реципиентов в нужном нам направлении. Так в Канаде, в Торонто мы таким путем вызвали регенерацию поджелудочной железы у крыс in situ на расстоянии около 20 км, но об этом чуть позже. Повторяю, мы разработали не просто новую технологию работы с одним из сотен типов лазеров, но создали квантовый биокомпьютер, имитирующий функции лазерных и радиоволновых излучений хромосом живых организмов. Мы используем лазер в красном диапазоне, а если брать ультрафиолетовый, то биокомпьютер будет на порядки мощнее. Здесь я заканчиваю физико-биологическую часть и перехожу в часть чисто биологическую, точнее генетическую.
Есть такое интеллектуальное достижение, которое называется моделью триплетного генетического белкового кода – триумф лауреатов Нобелевской премии Уотсона, Крика, а также Ниренберга и др. западных ученых. Основной вклад в создании модели белкового кода принадлежат Френсису Крику. Он, будучи гениальным человеком, однако, очень скромно оценил свою модель как генетический, ДНК-РНК-овый, способ шифровки последовательностей аминокислот в первичной структуре белков. Но эта модель была быстро и преждевременно превращена во всеобъемлющий генетический и одновременно в торговый брэнд. Может быть, на первых этапах это и было правильно. Полившиеся финансовые потоки в эту область биологии резко ускорили некоторое, более полное, понимание функций генетического аппарата. Почему я заговорил о модели генетического кода, ставшей на десятилетия каноном, не подлежащем критике? Каноны и догмы хороши в религии, но не в науке. Поэтому постепенно модель Ф.Крика из достижения превратилась в тормоз и систему отмывки денег "по научному".
В модели кода есть одно ключевое противоречие, которое Крик в 1953г. пытался снять с помощью так называемой "Вобл гипотезы". Она постулирует неоднозначные соответствия кодонов аминокислотам в ген-кодируемых белках и говорит о возможности не канонического и случайного спаривания первого нуклеотида антикодона транспортной РНК (тРНК) с третьим нуклеотидом кодона информационной РНК (иРНК) при ее трансляции в белок [Обозначения: 1-й, 2-й и 3-й нуклеотиды в кодонах и антикодонах – это для упрощения понимания. Тогда 1-й, 2-й и 3-й нуклеотиды в кодонах спариваются с 3-м, 2-м и 1-м нуклеотидами в антикодонах с учетом антипараллельности триплетов. Если быть точным с точки зрения химии полинуклеотидов, то 3-й "воблирующий" нуклеотид в кодоне – это так называемый 3’- нуклеотид, а комплементарный ему 1-й нуклеотид антикодона – это 5’- нуклеотид.]. Проще говоря, при биосинтезе белков иногда реализуется возможность нестрогого соответствия кодон-антикодоновых нуклеотидов в этом положении. Это значит, что образуются неканонические пары оснований [Каноническое термодинамически выгодное спаривание оснований – Аденин/Тимин, Гуанин/Цитозин (для ДНК), Аденин/Урацил, Гуанин/Цитозин (для РНК)], не отличающиеся существенно по геометрическим параметрам (Гуанин-Уридин и др). Кроме того, из Вобл-Гипотезы, да и просто из общей Криковской схемы (модели) кода, автоматически следует, что в кодонах (триплетах) генов только первые два нуклеотида (дублет) кодируют последовательности аминокислот в белковых цепях. Третьи же кодоновые нуклеотиды, не участвуют в кодировке аминокислотных последовательностей в белках. Эти третьи, хотя и детерминированы жестко молекулой ДНК, но допускают произвольные, случайные, не канонические спаривания с первыми нуклеотидами антикодонов транспортных РНК, переносящих аминокислоты. А посему эти первые нуклеотиды антикодонов могут быть любыми из 4-х возможных. Следовательно, третьи нуклеотиды в кодонах и спаривающиеся с ними первые в антикодонах, не имеют знакового характера и играют роль "стерических костылей", заполняющих "пустое место" в кодон-антикодоновых связках. Короче говоря, первый нуклеотид в антикодонах случаен, "воблирует" - от английского ‘wobble’ (качание, колебание, виляние). Это очень важно и скажу почему. Поскольку всего 64 кодона, а аминокислот 20, то получается избыток кодонов, и соответственно кодирующих дублетов. Этот избыток, используя лингвистическую аналогию, называют синонимией, то есть несколько дублетов кодируют одну и ту же аминокислоту, отсюда понятно существование изоакцепторных транспортных РНК (тРНК), переносящих аминокислоты на А-сайт рибосом. Такая синонимическая щедрость даже хороша – это избыточность информации. Но при белковом синтезе неизбежно возникают ситуации, когда одинаковые дублеты в кодонах информационной РНК, вынуждены как-то "кодировать" разные аминокислоты и/или стоп кодоны. Напомню, что третьи и первые нуклеотиды в кодон-антикодоновой "костыльной" связке находятся вне знаковой "игры", а это порождает лингвистическую неоднозначность, то есть ситуацию омонимии. Как, например, для слова "коса", когда смысл его неоднозначен. И соответственно, перед рибосомой, читающей одинаковые (омонимичные) дублеты кодонов, встает задача выбора из двух разных аминокислот, переносимых тРНК, одной единственной "правильной" аминокислоты и/или стоп-сигнала. Если выбор неправилен – белок получится аномальным, что может привести к метаболической катастрофе и гибели организма. Как же происходит правильный выбор? А он, как установлено, правильный и на 99,999% точен. Рибосома практически не ошибается, а казалось, должна бы, если автоматически следовать модели кода! Модель Ф.Крика это не объясняет. И сам Ф.Крик это видел и признавал в своих, опубликованных незадолго до смерти, воспоминаниях, но ограничился фиговым листочком Вобл Гипотезы. И вся последующая генетика никак не комментировала эту зияющую дыру в модели кода: модель противоречива, не точна, но синтез белков, тем не менее, безошибочен.
И только в 1978г. швед Ульф Лагерквист первым открыто заявил об этом противоречии в модели кода [Lagerkvist U., 1978, “Two out of Three”: an alternative method for codon reading. Proc. Natl. Acad. Sci. USA, v.75, p. 1759-1762.]. Но и Лагерквист также не предложил ничего внятного в объяснение, не обострил проблему, а лишь констатировал ее. Всех устраивало, что синтез белков – точный процесс и требовать от Криковской модели большего, вроде как, и не требовалось. "Жена цезаря вне подозрений". А потом появились работы о так называемых контекстных ориентациях при трансляции иРНК рибосомами. Это был момент истины. Было экспериментально показано, что выбор аминокислот рибосомой в омонимичных и других ситуациях зависит не только от дублетов кодонов, но и от контекста иРНК. Точно как в человеческой речи (текстах), когда, например, упомянутый омоним "коса" обретает точный смысл только в контексте целой фразы или предложения. Сей фундаментальный факт биологи опять-таки только констатировали. И все. Никакого анализа, почему так происходит и что это значит.
Меня такое положение дел не устраивало. Думаю, и многих других биологов. Это заговор молчания научного официоза. А причина проста. Анализ поведения генетического аппарата в ситуации омонимий логически приводит к сильному, я бы сказал, фундаментальному тезису о квази разумности генома. Для правильного выбора аминокислоты в ситуации омонимической неоднозначности белок синтезирующей системе необходимо ПРОЧИТАТЬ, прочитать не в метафорическом смысле, но реально, всю молекулу иРНК, то есть контекст и ПОНЯТЬ его смысл. И ПРИНЯТЬ РЕШЕНИЕ о выборе "правильной" аминокислоты или стоп-сигнала. Это можно сделать только С ПОМОЩЬЮ АКТА МЫШЛЕНИЯ, РАЗУМНО, что означает наличие у генома МЕНТАЛЬНОГО НАЧАЛА. Следовательно, геном базируется не только на чистой биохимии и физике взаимодействий информационных биомакромолекул, что стыдливо умалчивалось официальным научным сообществом в угоду жёсткому материализму и меркантильным соображениям. Понятие квази разумности генома выводит в совершенно другие области знания, которые сейчас пока либо недоступны, либо намеренно прикрываются. Одновременно это и пересмотр мировоззренческих позиций в отношении функции и происхождения наших хромосом, а заодно, пересмотр всех технологий работы генетиков, медиков. Следовательно, это ведет к переориентировке финансовых потоков в биологии и медицине, что многим не по вкусу. В дополнение скажу, что недавно в работах В.И.Щербака доказан ещё один фундаментальный факт – белковый код оперирует математикой с использованием запредельно абстрактного понятия НУЛЯ [Shcherbak V.I., 2003, Arithmetic inside the universal genetic code. BioSystems, v.70, pp.187–209.]. В.И.Щербак обнаружил в системе генетического исчисления использование функций ноля. Это чрезвычайно важное обстоятельство, поскольку ноль – сугубо мыслительное порождение, дающее начало координатному сознанию с его количественными мерами оценки внешнего мира, которые интерпретируется внутренним организменным генетическим сознанием-исчислением. Таким образом, цифры (наряду с буквами) становятся неотъемлемой частью генетического (белкового) кода. И мы должны принять очередное сильное положение, что in vivo существует арифметическое управление в лингвистической и/или текстовой генетике.
Итак, белок синтезирующая система и геном в целом обладают способностью к квази мышлению и математическому исчислению как части его. Замечу, что это не мышление уровня головного мозга, а квази мышление генетического аппарата как биокомпьютера. Здесь мы имеем дело с разными фрактальными размерностями одного и того же феномена – мышление и сознание на уровне коры головного мозга (одна размерность высшего порядка) и мышление и сознание на уровне функций хромосом и белок синтезирующей системы (другая размерность того же, но микро масштаба). Эти простые соображения выводят биологию, генетику, медицину в совершенно другие ареалы, и нам ясно, что каноническая модель генетического кода явно НЕ ПОЛНА. Геном – это лингвистико-математическая структура, тексты и алгоритмы Творца. При этом надо учесть и наши последние теоретико-экспериментальные разработки. В них показано, что генетические тексты и иные образные знаковые структуры генома могут храниться в форме поляризационных голограмм в жидких кристаллах хромосом [Тертышный, Гаряев, 2007, Волновые генетические нанотехнологии управления биосистемами. Теория и эксперименты. Новые медицинские технологии, 2007, №7, с.49-64]. Этот информационный пул огромен и занимает большую часть генома – около 98%. Удивительно, эти 98% генетического материала не поняты официальной генетикой и названы "мусором"! Но это тема для отдельного большого разговора.
Существенно также и то, что высшие формы сознания человека на уровне коры головного мозга реализуются, видимо, также хромосомным аппаратом, как основной информационной фигурой любых клеток. Коррелятом мышления и сознания опять-таки выступают текстовые структуры быстро синтезирующихся фракций белков. Замечу, что самый мощный синтез белков имеет головной мозг, нейроны коры мозга. Работа генома мозга идет также по пути голографирования с использование ресурсов ДНК. Таким образом, идеальное начало мышления-сознания базируется на материальных структурах информационных молекул ДНК, РНК и Белков, которые являются отображениями речи и технологий Творца, как инобытие Его Мысли.
Вернёмся к официальной генетике. Мы видим с её стороны полное игнорирование очевидных зияющих дыр в понимании работы генома, следовательно, и организма человека, животных, растений, бактерий, вирусов. К чему это ведёт и уже привело? Взять, к примеру, многомиллиарднодолларовую международную программу Геном человека. За теоретическую основу её взята Криковская модель генетического кода. Итог 15-летней программы таков – изучен порядок следования 3 миллиардов нуклеотидов в ДНК хромосомного набора человека. Найдено около 30000 генов. Это около 2% от всей ДНК генома человека. Остальная, главная часть, то есть эти 98%, научный официоз назвал мусором! Установлено, по этим генам человек практически неотличим от собственной кишечной палочки, от ослов, свиней, тем более обезьян. В основном это гены ферментов, которые, как рабочий инструментарий метаболизма, универсален почти для всех живых существ. Извините, господа генетики, я не согласен. Генетически мы существенно иные по сравнению с бактериями и свиньями! И отличия эти в иных принципах генетического кодирования, которые мы, и не только мы, обнаружили, и которые реализуются в тех самых 98% "мусора" геномной ДНК. А вы, господа официальные генетики и молекулярные биологи, об этом знать не хотите и объявляете лженаукой. Ну не вредительство ли? Программа Геном Человека впустую потратила огромные деньги, фактически отмыла их, обеспечив на многие годы безбедной жизнью кланы паразитирующих на незнании реальных псевдоученых. Но это полбеды. Хуже другое – неправильное, неполное понимание принципов генетического кодирования привело нас к проклятию генной "инженерии", когда тиражируются вирусные, бактериальные, растительные, животные химерные монстры, грозящие генетическим коллапсом всей планеты.
Что же предлагаем мы, пытаясь найти научные тропы к таким ясновидцам России, как Грабовой, Петров, Арепьев и их ученики? И что нас объединяет с ними? ДНК – это речь Творца и построены тексты ДНК сходно с человеческой речью. Много работ на эту тему опубликовано, особенно израильтяне много сделали. Это относится не только к известным генам, но мы полагаем, что голограммы генома в тех самых пресловутых 98% "мусора" содержат огромное количество интерференционно запакованных генов. Вот вам пример, косвенно подтверждающий эту мысль. Недавно опубликована статья группы Прюита из Соединенных Штатов [Lolle S.J., Victor J.L., Young J.M., Pruitt R.E. Genome-wide non-mendelian inheritance of extra genomic information in Arabidopsis. Nature, 434, 505-509 (2005).], причем не где-нибудь, а в Nature – нобелевский журнал! Так эта работа заставила вздрогнуть всю официальную менделевскую генетику. Как показали изящные эксперименты, скромное растеньице, ‘Резушка Таля’ (Arabidopsis thaliana), способно "вспомнить" и выставить напоказ предковый ген HotHead, который отсутствовал в его геноме. При этом HotHead (нормальный ген) заменял мутантный hothead. Полезная и многообещающая эндогенная манипуляция… Геном исходных мутантных семян предварительно анализировали блоттингом и секвенированием. Не было гена, а во взрослых растениях из этих семян ген HotHead выскакивал, как чёрт из коробочки! Откуда он взялся? Генетика до сих пор чихает от этого результата, а Мендель, наверное, перевернулся в гробу. Шум в Интернете среди научного официоза стоит страшный. Кстати, мы аналогичный результат ухода от мутаций получили за 2 года до американцев и опубликовали, но, увы, не в Nature, а посему он не замечен [П.П.Гаряев, Е.А.Леонова, 2003, Странный мир волновой генетики. Журнал "Сознание и физическая реальность", т.8, №6, с.27-40.]. Объяснение этому феномену простое – предковые гены исчезают, но не бесследно, они остаются в памяти генома, упакованные в голограммы "мусорной" ДНК. Если такие гены полезны организму, он их извлекает. Такие извлечения бывают аномальными и называются атавизмами. Однако я отвлекся. Так что же нас объединяет с ясновидцами, лечащими болящих СЛОВОМ и собственными излучениями? Связь очевидна – СЛОВО ЕСТЬ КВАЗИГЕНЕТИЧЕСКИЙ МАТЕРИАЛ на уровне микро- и макросоциальных отношений. Словом и лечат, и убивают. Разновидность такого рода воздействий на людей достаточно хорошо известна. Это нейро-лингвистическое программирование (НЛП). Кто только им не пользуется – от нашей мамы в детстве до таких говорунов как Жириновский. Но если лечащий, кроме искусства слова, владеет еще и повышенной по сравнению с нормой способностью излучать знаковые физические поля, то есть те, что излучают хромосомы, клетки и голова человека, то эффекты могут быть весьма значительными. Что мы и видим, начиная от воскрешения Христом Лазаря, кончая регенерацией органов и тканей у больных, вызываемые кудесниками ясновидения.
Излучательная часть феноменов такого рода особенно близка нам. Мы исследуем генетический аппарат не только как текстовую структуру, но как структуру, излучающую реальные, управляющие организмом физические поля – звук, свет, радиоволны. Этими полями мы научились оперировать как лечебными.
Одним из спектров электромагнитных полей, излучаемых хромосомами, точнее, их ДНК, является лазерный свет от ультрафиолета до ИК диапазона. Фактически, это предсказал наш ученый А.Г.Гурвич в 20-х годах прошлого века. Он же автор теории биологического поля. Именно его исследования стимулировали работу моей группы, начиная с 1985 года.
Теорию А.Г.Гурвича предельно кратко можно выразить одной фразой – хромосомы имеют волновой эквивалент.
Мне посчастливилось в течение 10 лет, с 1987 по 1997 годы, поработать в Физическом Институте АН среди талантливых физиков-лазерщиков и теоретиков. Помню, под влиянием идей Гурвича, я предложил группе учёных ФИАНа сделать лазер на молекулах ДНК и на хромосомах. Встречено было скептически, но убедил, и такую работу мы сделали – получили эффект лазерной накачки ДНК и хромосом in vitro с помощью пикосекундного лазера на парах меди. Это было опубликовано в журнале "Квантовая Электроника" в 1996 г. [А.М. Агальцов, П.П. Гаряев, В.С. Горелик, И.А. Рахматуллаев, В.А. Щеглов, 1996, Двухфотонно-возбуждаемая люминесценция в генетических структурах. Квантовая электроника, v.23, N2, с.181-184.] А спустя 6 лет близкие результаты получили японские авторы [Y. Kawabe, L. Wang, T. Nakamura, and N. Ogata Thin-film lasers based on dye-deoxyribonucleic acid-lipid complexes Applied Physics Letters -- August 19, 2002 -- Volume 81, Issue 8, pp. 1372-1374.]. Теперь уже никто не мог возразить, что хромосомы могут работать как лазерный источник и при этом фотоны его могут передавать генетический сигнал от клетки клетке.
Насчёт фотонной генетической передачи, это было гипотетично, хотя имелись кое-какие работы в этом направлении – исследования Дзян-Каньдженя, Казначеева, Бурлакова, Будаговского. Это наши, Россияне. Американцы тоже думали в этом направлении – Ричард Миллер в 1973 году первым предложил идею голографической памяти хромосом. Все это было, но не четко, без развитой теории и с малым запасом экспериментов. И вот в 1994 году мы совместно с Георгием Тертышным в Институте Проблем Управления РАН начали длительный цикл исследований, целью которых было создание аппаратуры, моделирующей знаковые (генетические) излучения хромосом в организме.
В 1996г. мне пришла в голову идея, что основной пул генетической информации записан в основном на уровне оптической активности ДНК хромосом в голографическом варианте, на способности вращать плоскость поляризации падающего на них света. Хромосомный континуум многоклеточного организма является поляризационной голограммой и одновременно лазером, считывающим генетическую световую информацию с самого себя. Чтобы доказать это, я предложил использовать и модифицировать специальный лазер, чувствительный к оптической активности хромосом и, соответственно, к информации, записанной в хромосомах на этом уровне.
Начали экспериментировать. Одна из серий опытов была особенно убедительна. Мы взяли чистую ДНК, выделенную из растения Arabidopssis thaliana. Препарат ДНК – это всегда испорченный генетический материал, это не целостные неповрежденные хромосомы. ДНК повреждается при выделении, удаляются специальные хромосомные белки. Вот такую испорченную ДНК-матрицу мы сканировали пучком нашего лазера. Такой тип лазера, как оказалось, осуществляет переход фотонов в радиоволны.
О переходе этом тоже особый длинный разговор. Мы дали физико-математический формализм этому явлению.
Эти радиоволны информационно связаны с фотонами лазерного луча. И если луч зондирует препарат ДНК, то информация, считанная с препарата ДНК, переписывается на радиоволны. А те, в свою очередь, переносят её на неопределенно большие расстояния во все четыре стороны. Вот мы и попробовали в 2000 году передать такую волновую генетическую и, напомню, поврежденную генетическую информацию с ДНК арабидопсиса на расстояние около 6 км между нашим Институтом Проблем Управления РАН и Институтом Общей Генетики РАН. Что из этого получилось?
А получилось то, что мы и ожидали. Проростки семян арабидопсиса-реципиента приняли передаваемую ложную информацию и мутировали. Сотни проростков мутировали. Чисто фенотипически это были так называемые летальные эмбриональные мутации по Мюллеру. Но мутации необычные, не связанными с разрывами ДНК в хромосомах реципиентов (энергия радиоволн мала). Во 2-м поколении мутации наследовались, а в 3-м ушли. Что это были за необычные мутации, вернее, квазимутации, нам до сих пор неясно. Вероятно, они были связаны с наведенными изменениями топологии хромосом растений, хромосом как динамичных жидких кристаллов (голограмм). Это был уход от мутаций и возврат к исходному фенотипу растений, аналогично тому, что наблюдала группа Прюита с геном HotHead, о чем уже говорил. Потом аналогичный квазимутагенез и уход от него в 3-м поколении мы увидели на растении картофеля. Подчеркну адресность воздействия – только близко родственные реципиенты воспринимали волновой морфогенетический сигнал.
Что означали такие, казалось бы, нехорошие эксперименты? Ведь мы повреждали геном реципиентов. Он подсказал нам направление будущих лечебных и прочих полезных воздействий на генетический аппарат такими радиоволнами. Если мы вызвали подобие неких мутаций у пространственно далеко расположенного реципиента, то нельзя ли таким образом избирательно уничтожать сорняки, насекомых-вредителей и переносчиков болезней, патогенные бактерии и вирусы и так далее, и тому подобное. Открылись широкие горизонты.
Любая хорошая идея должна быть простой в своей основе. Наша идея очень простая, но обросла гигантским физико-математическим аппаратом. Когда вы возьмёте наши статьи, поймёте, что не так всё просто, хотя основа ясна. Эта основа называется "делай как я". Если вы даёте волновую искажённую генетико-метаболическую информацию на объект воздействия, на реципиент, то получаете фактически адресное управление метаболизмом.
Сейчас о другом. Регенерация органов и тканей – то, что является главным в работе Аркадия Наумовича Петрова и то, что делают другие кудесники России. А нельзя ли нам регенерировать органы и ткани у людей нашей технологией? Это было бы просто здорово и подвинуло бы медицину в этом направлении, а также поддержало бы лечащих ясновидцев с позиции науки. И было бы великолепным подтверждением, что ментальные технологии ясновидцев базируются не на бреде сивой кобылы в лунную ночь, а основаны на классической физике и волновой генетике. Решили попробовать.
Есть так называемая аллоксановая модель диабета. Она достаточно изучена. Хорошо известно, что в методы лечения диабета вложены миллиарды долларов. Толку чуть. Инсулин – основное средство. А мы решили попытаться вырастить поджелудочную железу прямо в организме, использовав аллоксановую модель. Есть такой мощный яд – аллоксан. Достаточно крысе инъецировать 50 мг аллоксана, она гибнет от диабета – разрушаются бета клетки и инсулин исчезает из крови, что ведёт к гибели животных.
Первый эксперимент поставили в ИПУ РАН в 2000 году совместно с Институтом Теоретической и Эксперимнтальной Биологии в Пущино. Эксперимент был удачным, крысы выздоровели, поджелудочная железа регенерировала. Но крыс было мало – 36, поэтому со статистикой не проходило. Однако мы были замечены в Канаде и получили приглашение поработать именно по этой теме. Прилетели в Торонто и с 2001 по 2002 год работали в мощной фирме SynX. Там были идеальные условия для работы. Мы повторили наши положительные результаты на существенно большем числе крыс с хорошей статистикой. При этом продемонстрировали, что расстояние не играет критической роли для передачи волнового морфогенетического сигнала, способного запустить процессы регенерации поджелудочной железы in situ – прямо в теле животного. Расстояния от лазера до больных животных составляло от 1 см до 20 км. Результаты вызвали шок у канадских исследователей, которые участвовали в экспериментах. Эксперименты для нас были мгновенно свернуты. Нам сказали, что выздоровевших крыс усыпили! На вопрос – почему? Ответ был – эксперименты закончены. После этого мы вернулись в Москву, сами понимаете, в каком состоянии. Наши попытки возобновить эти работы в столице не встретили никакой поддержки, более того, академик Кругляков публично объявил меня лжеучёным, приводя в пример "бредовость" наших идей относительно создания лазера на молекулах ДНК. Я ему ответил открытым письмом в Интернете. А публикация японцев, подтвердивших наши результаты по ДНК-лазеру, поставила жирную точку на Круглякове. Словом, с 2002 по 2007 годы я пытался продолжить исследования. Безуспешно. А в ИПУ РАН меня уже не пустили, спасибо Круглякову.
Но судьба вывела меня на Аркадия Наумовича Петрова. Начались поездки по городам России, выступления, где я обосновывал научную состоятельность направления работ групп Грабового и Петрова, а заодно рассказывал о наших исследованиях. И нашлась-таки группа медиков, среагировали, попросили их обучить. Я им всё рассказал, обучил. Эти ребята полностью повторили канадские эксперименты, это опубликовано в ВАК-овском журнале [Гаряев П.П., Кокая А.А., Мухина И.В., Леонова-Гаряева Е.А., Кокая Н.Г., Влияние модулированного биоструктурами электромагнитного излучения на течение аллоксанового сахарного диабета у крыс. Бюллетень Экспериментальной Биологии и Медицины, №2, с.155-158 (2007)]. Удивительно, официальная наука позволила публикацию по волновым генам. Значит, какая-то подвижка имеется. Вслед за этой статьей опубликовали большой теоретический анализ полученных результатов [Тертышный Г.Г. Гаряев П.П., 2007, Волновые генетические нанотехнологии управления биосистемами. Теория и эксперименты. Новые медицинские технологии, 2007, №7, с.49-64], и одна в продолжение этого анализа еще предстоит.
ВОПРОСЫ
– Когда выйдет Ваша следующая третья книга?
П. Гаряев: Она практически готова, последние штрихи остались.
– У нас в Алма-Ате съедается червями яблоневый сад, то есть эту проблему тоже можно решить?
П. Гаряев: Так же, как мы вызывали мутации у арабидопсиса и на картофеле. Нужен препарат клеточных ядер или ДНК этих червей. Далее мы модифицируем этот материал рентгеном или ультрафиолетом. Это и будет источник ложной волновой информации для червей, которая дестабилизирует нормальную работу их генома. Черви погибнут.
– Как Вы подошли к теории Аркадия Наумовича? Как вы стыкуетесь?
А. Петров: Да я поругался в Интернете в их адрес, а он услышал.
П. Гаряев: И правильно сделал, что ругался. Я увидел в Интернете его критический комментарий в отношении безопасности наших работ. Пришел к Аркадию Наумовичу, объяснил, постарался убедить, что работы по волновой генетике будут развиваться независимо от того, хотим мы этого или нет, а также высказал соображения, что именно эти работы могут дать реальное научное обоснование ясновидению, понимание, как геном человека отвечает на речевые и ментально-волновые воздействия, исходящие от ясновидящего. С этого всё и пошло в наших контактах.
А. Петров: А когда эти процессы видишь в режиме ясновидения, видишь процессы сжатия, компрессии ДНК информации, то становится ясно, что в ДНК есть те же ситуации, о которых Пётр Петрович говорил. И когда хромосомная ДНК высвобождает свою волновую информацию, то она работает как голограмма, давая речевые и иные образы. При этом часть ДНК может читаться напрямую, когда она, как жидкий кристалл, составлена плотной упаковкой её нитей-текстов. И такие тексты могут читаться не только вдоль ее цепочек, но и поперек, справа налево, слева направо, вверх, вниз, вообще многомерно, по всем направлениям. Это огромная информация. И она, конечно же, имеет цифровой эквивалент.
П. Гаряев: Это как кроссворд, только он многомерный.
А. Петров: Есть ещё очень интересная фаза – когда поэтические тексты начинают оттуда выходить. Когда достигаешь определённого погружения в собственное сознание, анализируя ДНК-континуум, оттуда выходит очень значимая, но поэтизированная информация, то есть она рифмованная. Оказывается, что и такая возможность есть. При этом мы знаем, что разные народы пишут по-разному (справа налево, слева направо и так далее), то есть получается, что письменность и структуры ДНК имеют некую общность. То есть каждый народ отрабатывает какую-то глобальную программу, которая имеет фиксацию на уровне ДНК. Пётр Петрович со стороны науки, а мы с другой стороны, видим одно и то же. И мы также давно работаем со стволовыми клетками, регенерация именно на основе стволовых клеток ведётся.
П. Гаряев: Какая сейчас перспектива возникает. Вот этот широкополосный электромагнитный спектр ДНК, что мы извлекаем из нее. С ним можно оперировать по определенным программам и достигать самых разных биологических эффектов. Только осторожно надо работать. Тут непочатый край работы. Это тоже будет своего рода волновая генная инженерия. Только мы не будем идти напролом, как идут сегодняшние трансгенные "инженеры". Разобраться сначала надо в работе генома, а уж после манипулировать им на благо людей и всего живого на Земле.
Спектры, что мы регистрируем, можно получать не только с ДНК, клеток и организмов. Любое вещество дает эти необычные и неизвестные ранее спектры. Вообще это новый тип динамической поляризационно-лазерно-радиоволновой спектроскопии. Мы тут в самом начале. Например, можно получить спектры камней. Многие из них обладают биологической активностью, а спектры усиливают её во много крат. И все они дают свои уникальные мелодии, если перевести спектр из радиоволн в звук.
А. Петров: В своё время, нам привели мальчика, когда мы с Игорем Витальевичем Арепьевым работали, и у него была кризисная ситуация. И одновременно было видно, есть прогнозная фаза по будущему, что если сейчас выводишь ребёнка из этого кризиса, он дальше становится серийным убийцей, маньяком. И вот здесь разница между прибором и специалистом, который всё-таки развивает себя, а не прибор. Потому что он тогда имеет возможность прогнозировать будущее и знать, что произойдёт дальше. И по мальчику было найдено решение – сначала его определённое духовное развитие, и после этого – коррекция его здоровья. И тогда вот эта тяжелая прогнозная фаза ушла.
П. Гаряев: Подводя итог, вернемся к нашим экспериментам с крысами. Фактически, мы программировали стволовые клетки. Эта задача сейчас стоит перед медициной, поскольку программирование стволовых клеток – это возможность заменять старые и больные органы у людей на новые, затормозить их старение, продлить активную жизнь. Сюда вкладывают во всем мире миллиарды долларов. И, надо сказать, впустую. Без понимания принципов волновой генетики эта проблема станет черной дырой, куда впустую будут вбрасывать деньги и труд исследователей. Вопрос только – зачем? Ведь решение проблемы – вот оно, преподносится на блюдечке. Но это блюдечко тяжело далось – это результат нашей 20-летней теоретической и экспериментальной работы.
Приношу благодарность моим многочисленным соавторам и коллегам, с которыми я эти десятилетия работал, и без сотрудничества с которыми ничего бы не получилось. Особенно я благодарен физикам Оптического Отдела ФИАНа, с которыми мне удалось плодотворно поработать на протяжении 10 лет, а также моему наиболее близкому коллеге, блестящему физику – Георгию Георгиевичу Тертышному.
www.trinitas.ru
Критика ОТО
2012-04-30 02:03:00 (читать в оригинале)Введение
В предыдущей главе была доказана логическая противоречивость кинематики специальной теории относительности (СТО). Это заставляет вернуться к классическим понятиям пространства и времени. Поскольку релятивисты заявляют, что СТО является предельным случаем общей теории относительности (ОТО) в отсутствие гравитации, то сразу же возникают сомнения и в справедливости кинематики ОТО. В отличие от СТО ОТО содержит довольно интересные идеи, например, принцип эквивалентности выраженный через идею "геометризации". (Заметим, что неверность геометризации электромагнитных полей сразу очевидна: опыт показывает, что нейтральные частицы не чувствуют "электромагнитное искривление пространства".) Если бы базис ОТО был верен, ОТО могла бы претендовать на статус научной гипотезы о поправках к статическому закону тяготения Ньютона. Однако это не так и теория гравитации должна строится на другой основе. Справедливости ради надо отметить, что ОТО, в отличие от СТО, никогда не была общепризнанной безальтернативной теорией. Поток справедливой критики этой теории не прекращался с самого начала ее возникновения. Существует несколько хорошо продвинутых альтернативных теорий (например, [11,18]). Хотя мы не будем анализировать кроме ОТО иные теории, следует заметить, что теории, "играющие" в изменение свойств пространства и времени и имеющие своим предельным случаем релятивистскую кинематику СТО уже очевидно сомнительны. Основная цель настоящей Главы 2 — критика базисных понятий ОТО. Здесь будет продемонстрирована логическая противоречивость понятий пространства и времени ОТО. В Главе 2 шаг за шагом показываются правдоподобно скрываемые ошибки и спорные моменты из учебников [3,17,39]. Кроме общепризнанных интерпретаций ОТО мы будем также рассматривать некоторые "релятивистские альтернативы", чтобы прикрыть возможные лазейки для спасения этой теории. Обсуждаются вопросы синхронизации времени и принцип Маха, обращается внимание на сомнительные следствия из ОТО.
* * *
Многие трудности ОТО общеизвестны:1) нарушен принцип соответствия (без введения искусственных внешних условий не существует предельного перехода к случаю без гравитации);
2) отсутствуют законы сохранения;
3) относительность ускорений противоречит экспериментальным фактам (вращающиеся жидкости в космосе имеют форму эллипсоидов, в то время как невращающиеся — шара);
4) существуют сингулярные решения.
(Обычно, любая теория считается в подобных случаях неприменимой, но теория относительности для сохранения своего "всеобщего характера" начинает строить фантастические образы: черных дыр, Большого взрыва и т.д..)
* * *
Рассмотрим общие претензии к ОТО. Начнем с мифа "о необходимости ковариантности". Однозначное решение любого дифференциального уравнения определяется кроме формы уравнения еще заданием начальных и/или граничных условий. Если они не заданы, то в общем случае ковариантность либо ничего не определяет, либо при изменении характера решения может привести к физической бессмыслице. Если же задаются начальные и/или граничные условия, то при подстановке решений мы получаем тождества, которые и так останутся тождествами при любых правильных преобразованиях. Кроме того, для любого решения можно придумать уравнения, инвариантные относительно некоторого заданного преобразования, если определенным образом поменять начальные и/или граничные условия. Часто в ОТО используются аналогии с подпространствами, например, используют свернутый плоский лист. Однако, подпространство нельзя рассматривать отдельно от пространства в целом. Например, при свертывании листа в цилиндр обычно переходят для удобства в цилиндрическую систему координат, однако это математическое преобразование вовсе не влияет на реальное трехмерное пространство и реальное кратчайшее расстояние.
Простота аксиом и минимальность их количества еще не гарантируют правильность решения: даже доказать эквивалентность решений ОТО — трудная задача. Количество предпосылок, с одной стороны, должно быть достаточным для получения правильного однозначного решения, и, с другой стороны, должно обеспечивать широкие возможности выбора математических методов решения и сопоставления (у математики — свои законы). В ОТО, наряду с искусственным усложнением математических процедур, фактически введено дополнительное число "скрытых подгоночных параметров" из компонент метрического тензора. Так как реальное поле и метрика в ОТО неизвестны и требуют определения, то результат просто подгоняется под нужный с использованием малого числа реально разных опытных данных (вначале подглянули в ответ, а потом "с умным видом" считаем, что все так и должно быть в теории).
Если в СТО делалась хотя бы попытка экспериментально подтвердить постоянство скорости света и теоретически доказать равенство интервалов, то в ОТО даже таких попыток не сделано. Поскольку в ОТО в общем случае не имеет смысла , так как результат может зависеть от пути интегрирования, то могут не иметь смысла все интегральные величины и выкладки, использующие интегралы.
Множество вопросов заставляет сомневаться в правильности ОТО. Если общековариантность уравнений необходима и однозначна, то какой может быть предельный переход к классическим уравнениям, которые не общековариантны? В чем смысл гравитационных волн, если понятие энергии и ее плотности в ОТО не определено? И что (в отсутствие понятия энергии) выражает тогда групповая скорость света и конечность скорости передачи сигналов?
Степень общности законов сохранения не зависит от способа их получения (с помощью преобразований из физических законов или из симметрий теории). Получение интегральных величин и использование интегрирования по поверхности может приводить к иным результатам в случае движения поверхности (например, результат может зависеть от порядка предельных переходов). Отсутствие в ОТО законов сохранения энергии, импульса, момента количества движения и центра масс, которые подтверждены многочисленными экспериментами и работают на протяжении веков, заставляет весьма серьезно сомневаться в ОТО (следуя принципу непрерывности и преемственности развития науки). ОТО же пока еще ничем себя не зарекомендовала, кроме глобалистских претензий на принципиально экспериментально непроверяемую теорию эволюции Вселенной и нескольких весьма сомнительных подгонок под скудную экспериментальную базу.
Еще больше заставляет сомневаться в ОТО следующий факт: для одной и той же системы (причем только "островного" типа) с использованием вектора Киллинга иногда можно ввести некоторое подобие понятия энергии. Однако, при этом нужно пользоваться только линейными координатами, но нельзя, например, полярными. Не может же вспомогательный математический аппарат влиять на сущность одной и той же физической величины. И, наконец, нелокализуемость энергии и возможность "самопроизвольного" ее несохранения даже в масштабах Вселенной (это неприкрытый "вечный двигатель") заставляют полностью отказаться от ОТО и либо пересматривать концепцию "с нуля", либо использовать иные развиваемые подходы. Перейдем теперь от общих замечаний к более конкретным вопросам.
Вопрос о возможности изменении геометрии пространства в ОТО совершенно некорректен. Конечность скорости передачи взаимодействий может поменять только физические, а не математические законы. Не будем же мы утверждать, что прямая не существует, так как для ее проведения в бесконечность даже со скоростью света потребуется бесконечное время (аналогично для плоскости и пространства). Математический смысл производных тоже не может поменяться. Одна из демонстраций ОТО "о неизбежности изменения геометрии в неинерциальной системе" состоит в следующем: во вращающейся системе отсчета, вследствие сокращения длин, отношение длины окружности к ее диаметру будет меньше . Заметим, что никто не смог нарисовать для данного случая "новую геометрию": невозможно изобразить несуществующее. На самом деле не поменяется не только истинная, но даже наблюдаемая геометрия: не будет же математическая линия передвигаться или меняться при нашем движении. Хотя в теории относительности радиус, перпендикулярный движению окружности, меняться не должен, тем не менее предположим вначале, что окружность будет двигаться радиально.
* * *
Принцип эквивалентности
Принцип эквивалентности гравитации и ускорения может иметь отношение только к одной точке пространства, то есть нереален: это, например, уже приводило к неверному вычислению отклонения луча света в поле тяготения (только потом Эйнштейн подправил коэффициент в два раза). Принцип эквивалентности инертной и тяжелой масс в ОТО может быть строго сформулирован тоже только для одного отдельного тела (так как ОТО включает взаимосвязь пространства-времени и всех тел, то он нереален в ОТО). Поэтому физически ОТО не может иметь предельного перехода ни к одной нерелятивистской теории (а лишь формально математически). Все линейные преобразования СТО и ОТО относятся к пустому пространству, так как реальные тела (даже в качестве реперных точек) вносят нелинейности в свойства пространства. Поэтому различие явлений при переходе в другую систему отсчета должно изучаться строго в одной точке пространства и времени. Но как в одну точку поместить двух разных наблюдателей? Следовательно, все задачи СТО и ОТО могут носить только приближенный модельный характер (без глобализма). Нет ничего удивительного в том, что одна и та же величина — масса — может участвовать в разных явлениях: как мера инертности при воздействии любых сил, включая гравитационные, и как тяготеющая масса (например, движущийся заряд создает и электрическое и магнитное поле). Вопрос о точном равенстве гравитационной и инертной масс совершенно надуман, поскольку это равенство зависит от выбора численной величины гравитационной постоянной γ. Например, в случае пропорциональности mg = αmin все законы будут теми же, но с другим определением гравитационной постоянной γ' = α2γ. Не стоит искать здесь мистику и строить образы искривленного пространства. Подстановка одной и той же величины как для тяготеющей, так и для инертной масс производится не только в ОТО, но и в теории тяготения Ньютона. Просто это опытный факт (вернее, наиболее простой выбор величины ).
Когда говорят [37], что форма уравнений зависит от свойств пространства-времени, то в этом есть некоторая спекуляция. Создается впечатление, что мы как-то можем изменить это самое пространство-время для проверки данной зависимости. На самом деле мы имеем Вселенную в единственном числе. Попытка ОТО усложнить любое частное (локальное) явление добавлением сложности всей Вселенной не является позитивной для науки. Другое дело выбор локальных координат для математического описания локального явления (в этом случае конкретные симметрии явления упрощают описание), и глобализм опять ни при чем.
Использование неинерциальных систем в ОТО внутренне противоречиво. Действительно, во вращающейся системе достаточно удаленные объекты будут двигаться со скоростью, большей скорости света, а ведь СТО и ОТО утверждают, что видимые скорости должны быть меньше c. Однако, экспериментальный факт: фотография неба с вращающейся Земли показывает, что наблюдается видимое твердотельное вращение (классическое). Использование вращающейся системы (например, Земли) не противоречит классической физике при любом расстоянии объекта от центра, в то время как в ОТО величина компоненты становится g00 отрицательной, а это недопустимо в данной теории. Как же быть с наблюдениями в земной астрономии?
* * *
Понятие времени в ОТО также запутано до предела. Что же это за синхронизация часов, если она возможна только вдоль незамкнутых линий? Изменение момента начального отсчета времени при обходе по замкнутому пути — это явное противоречие ОТО, так как при большой скорости синхронизации можно сделать много подобных обходов и получить произвольное старение или омоложение. Например, представив вакуум (пустоту) вращающейся (если сами будем двигаться по кругу), мы можем получить разные результаты в зависимости от мысленного представления. Если на мгновение поверить в зависимость времени ОТО от гравитационного потенциала и эквивалентность гравитации и неинерциальности (ускорения), то легко понять, что тогда время зависело бы от относительного ускорения (расширенное толкование). Действительно, разным гравитационным потенциалам должны тогда соответствовать разные ускоренные движения и наоборот. Но относительное ускорение имеет векторный характер (и "спрятать" его невозможно), то есть расширенное толкование ОТО — единственно возможное. Используя модифицированный парадокс близнецов [51], легко доказать независимость времени от ускорения в расширенном толковании ОТО.
* * *
Некоторые следствия ОТО
Перейдем теперь к математическим методам ОТО и следствиям этой теории. Игры со свойствами пространства-времени приводят к тому, что в ОТО под вопросом оказывается применение вариационных методов: величины являются не аддитивными, преобразования Лоренца некоммутативны, интегральные величины зависят от пути интегрирования, даже не ясно как можно считать фиксированными конечные точки, если расстояния различны в разных системах отсчета.Нелокализуемость (неэкранируемость) гравитации приводит к тому, что в ОТО для наличия законов сохранения (только в системах островного типа) принципиально важны условия на бесконечности (евклидовость вследствие отсутствия масс на бесконечности) [37]. Классический подход более последователен и полезен в теоретическом и практическом приложении: энергия определена с точностью до постоянной, так как физический смысл имеет только локальное изменение энергии между двумя точками перехода. Следовательно, условия на бесконечности ни при чем.
Большое сомнение вызывает процедура линеаризации в общем виде, так как она может быть только индивидуальной. Говорится о стремлении к простоте, а даже времени вводят два типа: координатное и собственное. Часто производится подгонка под известный или интуитивный (классически) результат. Так один из знаков выбирается при расчете отклонения луча света, аналогично, для движения перигелия Меркурия [3] du/dφ может иметь два знака, какой выбрать? Не говоря уже о том, что производится деление на du/dφ, а эта величина может быть и нулевой. Пишется о сложности пространственно-временных связей, а в итоге очень долго переходят к привычным математическим координатам, иначе не с чем сопоставить результаты. За что же шла борьба? За наукообразие?
До сих пор нет достаточных экспериментальных доказательств того, какова скорость передачи гравитационных взаимодействий: больше, меньше или в точности равна скорости света (что постулируется в ОТО). Например, основываясь на данных наблюдений, Лаплас и Пуанкаре [24,87] считали, что скорость передачи гравитационных взаимодействий на несколько порядков превышает скорость света.
Теперь по-поводу экспериментального обоснования ОТО. Обычно, даже если есть сотня разных данных, не всегда строится теория — данные проще свести в таблицу. В случае же с ОТО мы имеем "Великую теорию трех с половиной наблюдений", из которых три — фикция. По поводу отклонения света в гравитационном поле от прямолинейного движения надо сказать следующее. Во-первых, как отмечало большинство экспериментаторов, количественное подтверждение эффекта существенно зависит от веры конкретного экспериментатора. Во-вторых, уже из классической формулы ma = γmMr/r3 следует, что любой объект, даже нулевой и отрицательной массы, будет падать в гравитационном поле. В-третьих, с чем, собственно, сравнивается эффект? С абсолютно пустым пространством? Еще в 1962 году группа Королевских астрономов заявила, что отклонение луча света вблизи Солнца не может рассматриваться как подтверждение ОТО, так как у Солнца существует атмосфера, простирающаяся на огромное расстояние.
Напомним, что явление рефракции учитывается для земной атмосферы астрономами уже очень давно. Еще Ломоносов обнаружил отклонение луча света в атмосфере Венеры. Для пояснения, представьте себе стеклянную сферу. Естественно, что параллельные лучи (от далеких звезд) будут отклоняться в ней к центру. Такая система всем знакома как оптическая линза. Подобная ситуация будет и для газовой сферы (атмосферы Солнца). Для точного расчета отклонения луча света в гравитационном поле нужно учесть наличие атмосферы Солнца и то, что наличие градиентов плотности и температуры на пути луча вызывает изменение показателя преломления среды и, следовательно, искривление луча света. И уж если на расстоянии сотни метров вблизи земной поверхности эти эффекты вызывают мираж, то не учитывать их для луча от звезды, проходящего вблизи Солнца миллионы километров — это чистая спекуляция.
Смещение перигелия Меркурия — эффект, конечно, красивый (но в единственном экземпляре — не мало ли для "привлечения научной теории"). Поэтому было бы интересно наблюдать его вблизи твердых тел (например, для спутников вблизи планет), чтобы можно было однозначно оценить его величину. Дело в том, что Солнце не является твердым телом и движение Меркурия может вызывать на Солнце приливную волну, которая может в свою очередь влиять на смещение перигелия Меркурия. (В зависимости от скорости передачи гравитационных взаимодействий и "гидродинамических" свойств Солнца волна может как опережать, так и отставать от движения Меркурия.) В любом случае необходимо знать скорость передачи гравитационных взаимодействий для вычисления влияния прилива от Меркурия и других планет на характеристики орбиты Меркурия, чтобы можно было отделить чисто "гравитационный" эффект общей теории относительности (если этот "чистый" эффект вообще существует).
При расчете в ОТО смещения перигелия (из строгого решения для единственной притягивающей точки) создается впечатление, что мы знаем точные массы астрономических тел. На самом деле, если мы пользуемся ОТО как поправкой к теории Ньютона, то ситуация противоположная: стоит задача по видимому движению планет восстановить их точные массы, чтобы потом подставить их для проверки ОТО. Представим себе, что орбита планеты — круговая. В этом случае сразу очевидно, что период вращения в теории Ньютона уже будет взят с учетом невидимой прецессии, то есть перенормирован. Поэтому в теорию Ньютона уже входят перенормированные массы. Поскольку поправки ОТО во много раз меньше возмущающего влияния всех планет и влияния несферичности, восстановление точных масс в этой сложной задаче многих тел может существенно изменить описание всей картины движения. Это нигде не учтено.
Вообще говоря, ситуация с описанием смещения перигелия Меркурия типична для поведения релятивистов. Во-первых, объявляется, что эффект был предсказан, хотя Эйнштейн сравнивал его с известными результатами приближенных расчетов Лапласа, полученными задолго до ОТО. Надеюсь, каждый человек понимает огромную разницу между "предсказать" и "объяснить задним числом" (вспомним анекдот от Фейнмана). Во-вторых, прецессия была и в классической физике: по данным 19 века итоговая величина прецессии за счет влияния некоторых других планет расчитывалась как 588'', а недостающая расчетная величина была всего около 43'', то есть составляла малую поправку. (Заметим, что по некоторым данным 20 века указывается общая величина прецессии почти на порядок большая, но при этом сохраняется из ОТО величина в 43'' — "табу"; впрочем, это может быть и опечатка — не будем придираться по мелочам к 1/3 от "огромной экспериментальной базы ОТО"). В-третьих, точный расчет в задаче многих тел даже современная математика пока выполнить не в состоянии.
В классическом случае расчет проводился как сумма независимых поправок от влияния отдельных планет (и Солнце и планеты считались материальными точками). Естественно, в классическом случае итоговый результат (уже более 90 процентов от наблюдаемого!) может быть еще улучшен при учете несферичности Солнца, влияния всех планет (и малых тел) Солнечной системы и того факта, что Солнце — не твердый объект (материальная точка) и его локальная плотность в разных слоях просто обязана "отслеживать" влияние остальных движущихся планет (на этом пути привлечения более реальных конкретных физических механизмов вполне может получиться недостающий малый эффект). Но то, что декларируют релятивисты — уму непостижимая спекуляция! Они "находят" эффект (причем только этот малый процент), рассмотрев движения лишь двух материальных точек — Солнца и Меркурия. Простите, а как Ваша ОТО подкорректирует уже найденную из классики большую часть эффекта? Боитесь считать? Тогда о каком "блестящем совпадении" вы твердите? Чистая подгонка под желаемое!
Прообраз "черной дыры" в решении Лапласа, когда свет, движущийся параллельно поверхности, начинает как искусственный спутник Земли двигаться по кругу, отличается от идей ОТО. Ничто не запрещает свету с достаточно большой энергией покинуть тело перпендикулярно его поверхности. Нет сомнения, что такие лучи будут существовать (и по внутренним причинам и по внешним): например, падающие извне лучи по закону сохранения энергии смогут набрать энергию и при отражении покинуть такую "черную дыру". Проще вместо привлечения противоречивых свойств света рассмотреть "падение" элементарной частицы, например, электрона. Остается ли для него возможность упругого отражения, или такую возможность (для спасения ОТО) нужно постулативно запретить? Если такую возможность все же не запрещать, то рассмотрим следующий процесс.
* * *
Даже из ОТО следует ненаблюдаемость "черных дыр": время образования "черной дыры" будет для нас, как отдаленных наблюдателей, бесконечным (даже если бы мы дождались "конца Света", ни одной "черной дыры" не успело бы образоваться). А поскольку коллапс не может закончиться, не имеют смысла решения, рассматривающие будто уже все произошло. Разделенность событий для внутреннего и внешнего наблюдателя бесконечным временем — это не "крайний пример относительности хода времени", а элементарное проявление противоречивости Шварцшильдовского решения. Этот же факт демонстрирует "неполнота" систем решений. Не ясно, что станет с законом сохранения заряда, если в "черную дыру" уйдет больше зарядов одного знака? Мистическое описание "метрических приливных сил" [39] при приближении к "черной дыре" неправомерно, так как это означало бы, что градиент гравитационной силы в пределах тела велик, а все идеи ОТО основаны на противоположных предположениях. Метрика Керра при наличии вращения также наглядно показывает несостоятельность ОТО: она математически строго дает несколько физически нереальных решений (те же операции, что и для Шварцшильдовской метрики не спасают положение). Таким образом, такой объект ОТО как "черные дыры" не может существовать и должен быть перенесен из сферы науки в область ненаучной фантастики. Вся Вселенная свидетельствует, что мир удивительно устойчив, часто динамически, но бесконечных коллапсов не бывает (скорее произойдет взрыв). Все это совершенно не отменяет возможность существования сверхмассивных (но динамически устойчивых) объектов, которые вполне могут проявляться рядом эффектов (например, аккрецией, излучением и др.). Для этого вовсе не нужны фантазии ОТО. Не нужно также искать пути искусственного спасения ОТО в виде "испарения черных дыр", так как такой возможности строго в ОТО просто нет (скорость света непреодолима), а вот в классике, напротив, нет никаких проблем.
ОТО содержит большое число сомнительных предпосылок и результатов. Перечислим некоторые из них. Например, требование для малых скоростей также и слабости гравитационного поля сомнительно: если посадить аппарат на массивную планету, неужели он не сможет стоять или медленно двигаться? Неужели, несмотря на температурные флуктуации не найдутся молекулы с малыми скоростями? Также рассмотрение центрально-симметричного поля в ОТО не имеет физического смысла: поскольку скорость может быть только радиальной, то не может быть не только вращений, но даже реальных температурных характеристик, то есть T = 0 K. Поле в полости не получают единым образом, а просто постулируют две разные константы, чтобы не было особенностей.
Излучение гравитационных волн для параболического движения (с эксцентриситетом ε = 1) приводит к бесконечной потере энергии и момента импульса, что явно противоречит опытным данным.
Фактически ОТО может применяться только при слабых полях и слабых вращениях, то есть в той же области, что и Ньютонова теория тяготения. Вспомним, что аналогичное взаимодействие между движущимися зарядами отличается от статического закона Кулона. Поэтому, прежде чем применять статический закон тяготения Ньютона, его надо проверить для движущихся тел, а это — прерогатива опыта.
Обсудим еще один принципиальный момент, касающийся относительности всех величин в ОТО. Законы, записанные просто как уравнения, сами по себе ничего не определяют. Для решения любой задачи нужно еще знание конкретики: характеристик тела (масса, форма и т.д.), начальных и/или граничных условий, характеристик сил (величина, направление, точки приложения и т.д.). Фактически задаются "реперные точки", относительно которых и изучаются последующие изменения величин (положения, скорости, ускорения и т.д.). Принципиальная относительность всех величин в ОТО противоречит опытам. Последующая искусственная попытка выводить ускорения (или вращения) относительно локальной геодезической инерциальной Лоренцовой системы — это просто подгонка к единственно работающим и экспериментально проверенным координатам абсолютного пространства (ОТО органически ничего подобного не содержит [18]).
Гравитационная постоянная не является математической константой, а испытывает вариации [9]. Следовательно, данная величина также может учитывать поправки к статическому закону тяготения Ньютона (например, не проведено анализа этих влияний при расчете смещения перигелия Меркурия). Напомним, что при финитном движении (например, периодическом) в связанной системе многих тел могут наблюдаться различные резонансные явления, выражающиеся в согласованных корректировках параметров орбит (особенно с учетом конечных размеров тел — несферичности их формы и/или распределения масс). Вообще говоря, принцип близкодействия может оказаться для гравитации полезным (а может и нет, в зависимости от скорости передачи гравитационных взаимодействий) только в ограниченном числе случаев: при быстрых (v → c) движениях массивных (одного порядка) тел вблизи друг друга. Автору неизвестны подобные практические примеры.
Подход ОТО к гравитации уникален: закрыться в кабине лифта, наслаждаясь падением, и не знать, что через мгновение расшибешься. Конечно, в реальности ситуация иная: мы всегда видим, куда и как мы движемся относительно притягивающего центра. Вопреки Тейлору и Уиллеру это и есть вторая "частица", вместе с наблюдателем — первой "частицей". Именно поэтому чисто геометрический подход к гравитации является временным ответвлением на пути физики (хотя, как расчетный инструмент может когда-нибудь оказаться полезным). И двум путешественникам в притче из книги [33] (якобы демонстрирующим подход геометрии искривленного пространства) нужно "совсем немного": желание двигаться от экватора именно вдоль меридианов по шаровой поверхности Земли, а у остальных пяти миллиардов человек может не оказаться такого желания. В отличие от желания путешественников, сколько бы вы не пожелали не притягиваться к Земле или Солнцу и без усилия улететь в космос, вашего желания явно недостаточно. Подобное явление и отражает понятие силы (в данном случае силы тяжести). Геометрия не может ответить на вопросы: сколько типов взаимодействия реализуется в природе, почему только они, почему существуют локализованные массы, заряды, частицы, почему сила тяжести пропорциональна именно второй степени расстояния, почему те или иные конкретные физические постоянные реализуются в природе и на многие другие. Эти вопросы — прерогатива физики.
Критика космологии
Теории эволюции Вселенной навсегда останутся гипотезами, так как ни одно из предположений (даже об изотропности и однородности) не может быть проверено: "давно ушедший и движущийся поезд можно догнать только в другом месте и в другое время". ОТО приписывает себе разрешение ряда парадоксов (гравитационного, фотометрического). Напомним, что гравитационный парадокс состоит в следующем: для бесконечной Вселенной равномерной плотности невозможно из уравнения Пуассона получить определенные значения для гравитационного ускорения тел. (Какое отношение к реальности имеют чисто математические неопределенности с условиями на бесконечности в физической модели?) Напомним также суть фотометрического парадокса: для бесконечно существующей (стационарной) бесконечной Вселенной без учета поглощения и преобразования света яркость неба должна равняться средней яркости звезд (опять много нереальных предположений). Однако и в классической физике были описаны возможности решения подобных парадоксов (например, с помощью систем разных порядков: сфер Эмдена, структур Шарлье и др.). Очевидно, что Вселенная не является размазанной средой и мы совершенно не знаем ее структуру в целом, чтобы утверждать о возможности реализации условий для подобных парадоксов (скорее, наоборот).
Например, фотометрический парадокс Ольберса легко понять на основе аналогии с океаном: свет поглощается, рассеивается и отражается порциями и на определенную глубину свет просто перестает проникать. Конечно, для разреженной Вселенной такая "глубина" огромна. Однако, светящиеся звезды представляют собой довольно компактные, далеко отстоящие друг от друга объекты. В результате в интенсивность света ночного неба вносит вклад лишь конечное число звезд (не говоря уже о том, что в теории надо учесть еще и эффект Допплера, а еще лучше — экспериментальный факт — красное смещение).
По-поводу красного смещения в спектрах астрономических объектов ситуация не до конца определенная. Во Вселенной существует значительная доля объектов, у которых разные участки спектра имеют совершенно различное смещение. Вообще говоря, поскольку расстояние до отдаленных объектов прямо не определяется (вычисляемый результат завязан на определенные гипотезы), то связывать его с красным смещением — тоже гипотеза (в которой неизвестно, что может быть проверено). Например, расширяющаяся Вселенная и без ОТО дает красное смещение согласно эффекту Допплера. Кроме того нужно учесть, что в красное смещение и наполнение так называемого реликтового излучения будет давать вклад элементарное рассеяние: вспомним, что эффект Комптона дает волны с λ' = λ0. Смещение линий в гравитационном поле прекрасно предсказывалось даже механистическими моделями из общих энергетических соображений.
Вообще говоря, теория Большого Взрыва вызывает Большие сомнения. Помимо банальных вопросов: что взорвалось, куда и когда (ведь не было ни пространства, ни времени, ни материи), возникает вопрос: а как же быть с выводами ОТО о черных дырах (непреодолимостью предельной скорости света)? Ведь Вселенная должна была быть в нулевой момент черной дырой (да и не только в этот момент, а в течение некоторого периода времени). Как же быть с ограничениями ОТО, ведь теперь вместо такого образного описания сжатия в черной дыре мы экспериментально наблюдаем повсеместное расширение? Интересно, наверное, сочинять то, что нельзя проверить (только не стоит называть это наукой).
Перейдем к следующему принципиальному вопросу. Является ли плюсом то, что распределение и движение материи не могут быть заданы произвольно? И правильно ли это? В общем случае это означает противоречивость теории, так как кроме гравитационных сил существуют и другие силы, способные перемещать материю. С практической точки зрения это означает, что мы должны были и в начальный момент времени задать все распределения "правильным для ОТО" образом. Тогда не к "моменту творения" ли мы должны относить ? И какие принципы должны быть однозначно детерминированными для такого выбора? Знаний требуется больше, чем любые возможные ожидания от предсказаний ОТО.
Под вопросом оказываются возможность точечного описания и теория возмущений, ведь итоговые величины тоже не могут быть произвольными. Присоединение к системе уравнений совершенно неизвестного уравнения состояния означает искусственное усложнение связью макро- и микроуровней и отражает возможность произвольных подгонок (например, выбрасывается температурная зависимость). Возможность добавления космологической постоянной в уравнения Эйнштейна — это косвенное признание неоднозначности уравнений ОТО и возможности произвола. Уж если с такой точностью все можно задавать, то почему произвольным образом не задавать первоначальное распределение и движение материи.
* * *
Принцип Маха обусловленности инертной массы и абсолютности ускорения действием далеких звезд также сомнителен, так как объясняет внутренние свойства одного тела через свойства других тел. Конечно, идея сама по себе красивая. Если считать, что все в мире взаимосвязано и существует некоторое идеальное полное уравнение состояния, то любое свойство тел должно определяться влиянием всей остальной Вселенной. Однако, тогда пришлось бы каждую частицу считать индивидуальной. Этот путь порочен для науки, идущей от меньшего знания к большему, так как "нельзя объять необъятное". Практически, если учесть неравномерное распределение массы (в компактных объектах) и разные величины сил притяжения от ближних и дальних объектов, то получилось бы сплошное "дерганье" вместо равномерного вращения или равномерного движения по инерции. Принципиально принцип Маха не может быть проверен: как удаление всех тел из Вселенной, так и искусственное устремление гравитационной постоянной к нулю - ничего не имеющие с реальностью абстракции. Однако, экспериментально можно оценить влияние "далеких звезд", считая массу Вселенной сосредоточенной в основном в компактных объектах.
* * *
Выводы к Главе 2Данная Глава 2 была посвящена критике ОТО. Здесь было выделено множество бросающихся в глаза сомнительных моментов из учебников по ОТО, начиная с общих положений о ковариантности, базовых физических понятиях и кончая более конкретными. Подробно проведено доказательство неизменности геометрии во вращающейся системе. Обсуждена необоснованность и противоречивость в ОТО принципа эквивалентности. Продемонстрирована противоречивость понятия времени и его синхронизации в ОТО. Для наиболее интересных частных случаев показаны способы синхронизации времени и одновременного измерения длин. В Главе 2 показывается неизменность геометрии пространства и обсуждается роль границ. Сомнительные моменты подчеркнуты как для методов, так и для многочисленных следствий ОТО. Подробно рассмотрены противоречивость понятия "черных дыр", Шварцшильдовского решения и многих других решений и следствий ОТО. Также обсужден принцип Маха и его возможная проверка.Итоговый вывод главы заключается в необходимости возврата к классическим понятиям пространства и времени и построении теории гравитации на этой прочной основе.
Дайджест главы 3. Эксперименты ТО
Введение
В предыдущих главах значительная часть критики теории относительности базировалась на так называемых мысленных экспериментах. Сделаем одно тривиальное замечание, чтобы у какого-нибудь "доброжелателя" случайно не возник абсурдный вопрос о технической осуществимос
Энергия нулевой точки
2012-04-29 20:58:00 (читать в оригинале)Итак что же такое энергия нулевой точки? Для начало стоит включить своё воображение и представить две некие равные силы которые противодействуют друг другу (школьная физика) в точке их противодействия энергия равна нулю, так как эти две силы компенсируют друг друга, Но при этом мы не можем утверждать что не производится никакой работы так как силы друг друга компенсируют!

Так как две эти силы затрачивают некоторую энергию на компенсацию другой равной силы. То есть общая энергия двух сил будет равна F1+F2. И так примерно разобрались что же представляет собой нулевая точка. Нулевая точка- это точка в которой достигается нулевое значение сил при противодействии двух сил.
Перейдём к более тонким материям. Итак что такое всемирный эфир? Эфир- это некая всепоглощающая субстанция которая присутствует везде и участвует в процессе энергетического обмена в элементарных частицах. К чему я завел речь о эфире? Эфир можно так сказать это тоже некая само уравнивающая субстанция которая представляет собой модель взаимокомпенсирующих сил , где так же содержится нулевая точка. Эфир обычно сравнивают с водой, погрузившись в воду, вода производит практически равномерное воздействие на тело, а именно давление. Можно представить некий пузырь погруженный в воду, на который со всех сторон действует сила уравниваясь в ноль, тем самым мы можем сказать что работа здесь не производится, как только стоит пузырю лопнуть, мы будем наблюдать работу масс воды и соответственно получим импульсную энергию, которая в течении некоторого времени будет совершать колебания в месте бывшего пузыря.
И так , как же будут происходить эти процессы в электродинамике. А вот здесь самое интересное!
Ниже находится изображение осциллограммы катушки которая находилась под действием кратковременного электрического импульса.
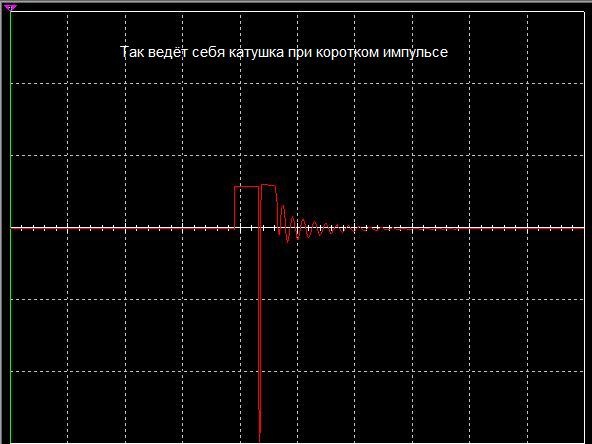
И так на изображении показана осциллограмма катушки индуктивностью 10 Гн без магнитного сердечника, слева импульс поданный на катушку амплитуда которого равна 5 вольт, далее мы видим процесс самоиндукции, который по длительности импульса полностью покрывает затраты на первичный импульс и даже больше производит энергии, чем при первоначальном импульсе.
Ниже находится изображение осциллограммы работающего устройства на самогенерации, то есть без внешних источников питания.
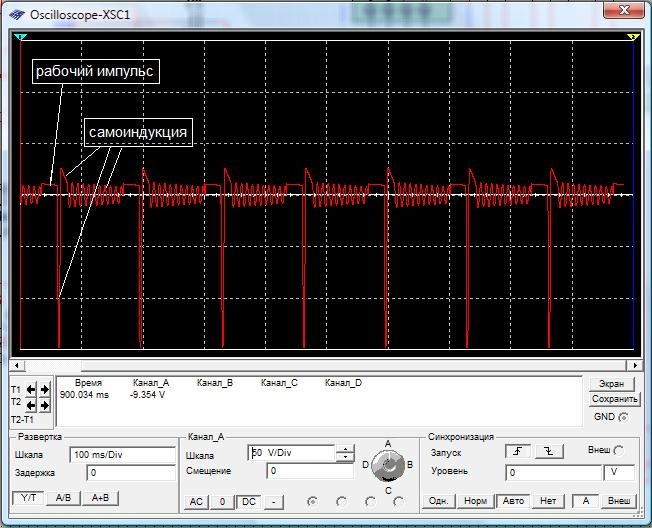
..Нужно правильно рассчитать длительность импульса и конечно использовать только крутой импульс прямоугольной формы, так же нужно учитывать время насыщения катушки и не переборщить с ним, чтобы не было дополнительных затрат мощности. Главное точный расчет! Можно поэкспериментировать и изменить схему домашних кварцевых часов , которые в своей работе используют катушку с большой индуктивностью и сделать из них вечные часы без использования батареек- вполне реально!
И так теория очень проста, при подачи импульса на катушку вокруг катушки образуется электромагнитное поле, что приводит к смещению эфира. После резкого пропадания поля (так как не используются сердечники в катушке) Происходит лавинное смещение эфира в начальное положение из-за чего и возникает импульс самоиндукции превышающий рабочий импульс! Внимание импульс должен быть крутой по своей форме! Аналогия или вы бросаете камень с высоты на землю или плавно кладете его на землю, в первом случае работа камня будет больше.
Вот один из способов использования энергии нулевой точки.
next-energy.2x2forum.ru
Энергия нулевой точки в данном случае это энергия эфира, который находится в уравненном состоянии. Когда происходит импульс в катушке мы выводим из равновесия эфир, смещаем его полем магнитного импульса. После исчезновения магнитного импульса эфир стремится занять исходное положение, но с более высокой скоростью, что и приводит к возникновению самоиндукции в катушке.
next-energy.ru
Энергия, которая попадает в катушку через маленькое энергетическое окошко (нулевая точка), может быть использована для производства любой работы, представляясь ключом к успеху этой системы. Здесь, в нулевой точке, энергия конденсируется в положительные и отрицательные компоненты тока. Когда энергия вытекает из «стока», магнитное поле обваливается и сильное магнитное потрясение создается следом за ней. Правильно настроенная система может захватить и преобразовать «лучевую энергию» описанным способом.
Н. Тесла
Магнитный компонент "окружающего фона" можно возбудить (вывести из равновесия) диполем или резонансным трансформатором (зависит от устройства), и что этот дисбаланс позволяет собрать большой объем электроэнергии, используя конденсаторы и катушки индуктивности. Во-вторых, можно отбирать одновременно столько «копий» выхода энергии, сколько нужно, от одного магнитного возмущения, без какого-либо его ослабления. Это позволяет получать намного больше мощности, чем затрачено на магнитное возбуждение.
Д. Смит
Влияние плотности эфира на скорость существования материи
2012-04-29 19:28:00 (читать в оригинале)Фролов А.В.
Генеральный Директор
ООО «Лаборатория Новых Технологий Фарадей»
office@faraday.ru
Экспериментальные данные от различных источников показывают, что скорость существования материи зависит от плотности эфира. Это вполне объяснимо, поскольку частицы материи являются вихревыми процессами в эфире. В таком случае, ход времени, имеющий физический смысл скорости существования материи в пространстве определяется параметрами вихря. Скорость хода времени, как и само понятие «время» может быть устранено из рассмотрения в физике, как вторичный термин. Первичным является понятие «скорость процесса».
Приведем примеры простейших конструкций, создающих изменения плотности эфира без применения источников энергии. Их работа основана на том, что планета находится в движении относительно эфира.
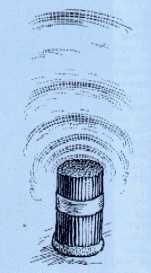
Пучок трубок создает эффект полостных структур (стоячие волны де Бройля). Открытие Гребенникова и Золотарева.
На Рис.1 показан пучок трубок в качестве простейшего излучателя. На Рис.2 показа спиральный излучатель, применяемый в медицине 19-го века.
Эти рисунки мы нашли в книге В.С.Гребенникова «Мой мир». На рис.3 показан конусный излучатель Гребенникова (внешний вид). Аппарат изготовлен из картона и включает в себя три постоянных магнита. Постоянные магниты в данном устройстве необходимы для возбуждения потоков эфира, поскольку магнитное поле можно рассматривать, как замкнутый циркулирующий поток эфира.
Рис.3
И наконец, рисунок Гребенникова, на котором изображен известный конструктивный элемент в форме пирамиды (Рис.4) позволяет понять, почему точки над вершиной и в центре пирамиды проявляют особенные свойства.
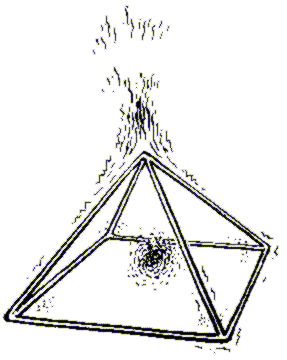
Рис.4
Ребра пирамиды создают поток эфира над ее вершиной, а в центре создается область уменьшения плотности эфира. Экспериментально показано, что помещенные в эти области материальные объекты изменяют свои физико-химические свойства.
Еще один пример данной технологии: открытие Богданова («Аргументы и факты», №10, 2002). Увлеченный идеей омоложения, автор из Башкирии не только придумал (увидел во сне), но и создал макет устройства, которое позволяет изменять свойства вещества, помещенного в него. Устройство состоит из сферического комплекса конусных излучателей, работа которых в роли отражателей эфира аналогична устройствам, показанным на Рис. 3 и Рис.4.
 Рис.5Капсула омоложения Богданова состоит из нескольких конусных элементов, образующих сферу с полостью в центре.
Рис.5Капсула омоложения Богданова состоит из нескольких конусных элементов, образующих сферу с полостью в центре.Макет был изготовлен из картона и склеен эпоксидным клеем. Диаметр макета составляет около 50 см. Реальное устройство Богданова должно иметь диаметр внешней сферы 30 метров и диаметр внутренней (пустой) сферы 8 метров. Проверка эффекта в одном из московских оборонных предприятий показала, что внутри макета наблюдается структурирование раствора марганцовки (кристаллы соединяются в шарик). Очевидно, что данная конструкция относится к области резонаторов эфирных волн (волн материи де Бройля) и позволяет намного усилить эффект за счет фокусировки.
Отметим, что применение пассивных отражателей и фокусировка потоков эфира является таким же примитивным методом изменения плотности эфира, как парусные технологии в сравнении с современными силовыми установками судов. Электромагнитные явления, которые хорошо изучены и широко применяются на практике в нашей жизни, могут быть заново осмыслены с позиций эфиродинамики, если мы будем искать способы активного воздействия на эфир. Такие попытки были в свое время предприняты В.А.Чернобров, Москва. На Рис. 6 показана схема установки, в которой создается управляемое влияние на плотность эфира.
Рис.6
В совместном проекте с Чернобров В.А. нами было построено и испытано устройство, показанное на Рис. 7.

Рис.7 Изучение эффекта в «Лаборатории Новых Технологий Фарадей»
Элемент данного устройства разработан Чернобров В.А. и имеет форму трех-витковой катушки, показанной на Рис.8. Мы подробно рассматривали данную технологию в статье «Способ и устройство управления темпоральными характеристиками физических процессов», журнал Новая Энергетика, №3 (12) 2003, стр. 4.
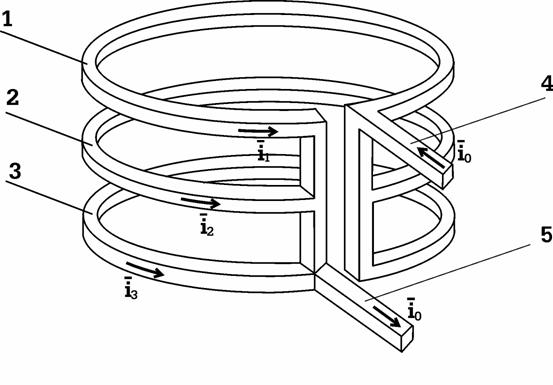
Рис.8
Помимо электромагнитных явлений, в качестве механизма управления плотностью эфира могут применяться любые необратимые процессы, идущие с изменением энтропии. Это было убедительно показано в работах Н.А.Козырева. Схема эксперимента на Рис. 9 предлагается для постановки совместного проекта с заинтересованными партнерами.
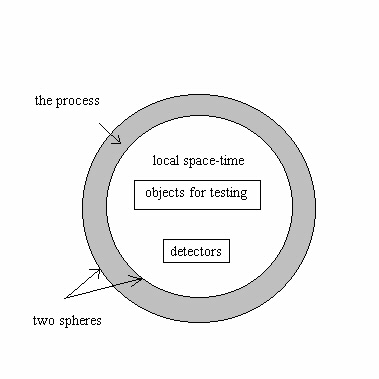
Рис.9
В роли процесса могут быть использованы фазовые переходы вещества из твердого в жидкое состояние или процессы кристаллизации. В данном эксперименте мы планируем доказать ранее полученный вывод о том, чтоуправляемое изменение плотности эфира возможно. В отличии от электромагнитных методов, применяемых в экспериментах с Чернобров В.А., в новом проекте исключается влияние электромагнитных полей на датчики и объекты в области фокусировки потока эфира.
Мы ищем партнеров и заинтересованных заказчиков для развития данной технологии.Прикладные аспекты управляемого изменения плотности эфира выходят далеко за рамки самых смелых фантазий о «машине времени», поскольку мы открываем дверь в мир управления параметрами существования любой материи. Это не только вещества с новыми свойствами. Для живой материи изменение условий ее существования означает возможность ускоренного развития, оптимизации деления клеток, усиление защитных свойств организма и раскрытие его новых качеств.
alexfrolov.narod.ru
Кто и зачем всучил науке СТО+ОТО?
2012-04-29 19:17:00 (читать в оригинале)В своё время евреев с Библией намеренно впихивали в первоисточники христианства. Подтасовывали археологию, сочиняли памятники, даже после Второй мировой войны дали землю под "исторический" Израиль… Однако наука стала отклоняться от нужного направления.
Понадобился мощный аккорд библейско-еврейской темы, исполненной по "математической" партитуре, расписанной финансовой олигархией. Обратите внимание, что все авторы-антиэйнштейновцы подробно проанализировали КТО и КАК протолкал Эйнштейна в гении, но вопрос ЗАЧЕМ, даже не затрагивали!! Симптомчик, однако... "А Ларчик просто открывался".
1) СТО привлечена для того, чтобы люди ни в коем случае не искали способы связи с "тонкими мирами" и не пытались "участвовать" в управлении космосом (мировым порядком). Для этого им навязали высосанные из пальца проблемы:
- как устроен некий эфир (греч. aither - (ясное) небо, верхние слои воздуха, (чистый) воздух < aitho - воспламеняю, горю, пылаю, сжигаю), призванный подменить физическое пространство;
- как справиться со множеством совершенно дурацких парадоксов, порожденных всё запутывающим лихим маневром, связавшим пространство и время в некий единый "пространственно-временной континуум", проще говоря, породив эдакую "кривеньку качечку" (укр. хромую уточку). Именно с неё началась релятивистская физика, а космология приобрела отчетливые черты МЧС (математической чуши собачьей).
2) ОТО привлечена для сохранения общей идеи о разовом творении Вселенной, о существовании её границ и как бы потустороннего мира. Человеческую мысль перенаправили на путь математического исследования:
- как устроен некий эмпирей (от греч. empyros - огненный, объятый пламенем), т.е. место за вселенским пространством;
- зачем создана Вселенная и доколе ей расширяться;
- когда и за что Вселенную ликвидируют. Кстати, ликвидация Вселенной является параллельным (подобным) процессом ликвидации человечества за нарушения (грехи) христианских заповедей.
Есть такая статья "Есть ли у Вселенной история?" (2010), где сказано: "XIX век примечателен тем, что "математические физики" вернулись к исходной идее древних - "сфере неподвижных звезд" (как бы её сейчас ни называли и в какие бы теоретические одежды ни рядили). В данном случае не имеет никакого значения, где находится центр Вселенной - на Земле, на Солнце или там, где мог бы оказаться выпихнутый с Земли наблюдатель. Главное - есть окраина мира, которая очень привлекательна для христианской идеологии, т.к. оставляет полным-полно места за пределами "звездной сферы" и для Бога, и для рая с адом, и для бизнесменов с наворованными "финансовыми средствами".
Уверен, что современным ученым физико-математических наук перечисленные задачи по плечу, а Нобелевский комитет своих не обидит".
Вне всякого сомнения всё упирается в "физико-математическое" доказательство основ христианства, как единой идеологии мирового сообщества алчных ПОТРЕБИТЕЛЕЙ (точнее идеологии для ширнармасс за вычетом "золотого миллиарда". См статью "Три религии в одной Библии", 2010).
Итак, когда подвернулся удобный момент, то нашелся звезд с неба не хватающий, но очень покладистый Эйнштейн, который подкинул пару кирпичиков в основы мироздания. Благо подвернулась "Воображаемая геометрия" Лобачевского. Ну, как тут не воспользоваться таким подарком? Вот вам и современный заменитель Моисея (или продолжатель дела?), хоть неряшливый и лохматый, но очень послушный.
Обратите внимание, еврофизику, выполнившему свою главную задачу, было позволено болтать довольно много. А чтоб не путался под ногами, для него даже создали "Институт высших исследований" (в 1933 г., г. Принстон, штат Нью-Джерси), где он проработал до конца жизни (1955).
Я уж не говорю, что докторскую ему присвоили в 1909 без защиты (так сказать "почетно"), нобелевку пытались впарить ежегодно 11 лет подряд (1910-1921), пока всё-таки не уговорили физиков согласиться. Но дали "за его заслуги в теоретической физике, и специально за его открытие закона фотоэлектрического эффекта". Справка: этот эффект был открыт в 1886 г. Г. Герцем, закон установлен в 1888 г. А.Г. Столетовым.
Думаю, потребность воздаяния псевдомессии по заслугам (выкинуть на политическую свалку истории) возникнет ещё не скоро. Не все мирские проблемы решены в соответствующую пользу!
В.А. Вейник, Письмо Л.А. Полыковскому, 10.01.2012.
veinik.ru
Категория «Радио»
Взлеты Топ 5
|
| ||
|
+40 |
47 |
Фрагменты |
|
+30 |
57 |
тот_самый_Петрович |
|
+19 |
40 |
история интерьера |
|
+1 |
27 |
Новости сайта RocketsMusic.ru |
|
+1 |
17 |
промо радио |
Падения Топ 5
|
| ||
|
-17 |
20 |
Радио ФМ Онлайн слушать бесплатно |
|
-24 |
2 |
Лучинин.net |
|
|
|
|
|
|
|
|
|
|
|
|
Популярные за сутки
Загрузка...
BlogRider.ru не имеет отношения к публикуемым в записях блогов материалам. Все записи
взяты из открытых общедоступных источников и являются собственностью их авторов.
взяты из открытых общедоступных источников и являются собственностью их авторов.